Article Text

Abstract
Introduction Obesity is a risk factor for type 2 diabetes mellitus (T2DM) and cardiovascular disease. T2DM increases the risk of cardiovascular-related death. We investigated changes in circulating exosomal microRNA (miRNA) profiles in patients with DM with obesity compared with patients without DM with obesity.
Research design and methods This prospective study involved 29 patients with obesity (patients without DM=16, patients with DM=13) and healthy volunteers (HVs) (n=18). We measured circulating levels of exosomal miRNAs by next-generation sequencing and compared miRNA levels across the three groups.
Results The expression levels of 25 miRNAs (upregulated=14, downregulated=11) differed between patients with obesity with DM and patients with obesity without DM. Compared with HV, patients with DM with obesity had 53 dysregulated miRNAs. Additionally, moving stepwise from HV to patients with obesity without DM to patients with obesity with DM, there was a consistent increase in expression levels of miR-23a-5p and miR-6087 and a consistent decrease in expressions levels of miR-6751-3p.
Conclusions Our data show that the exosomal miRNAs is altered by dysregulated glucose metabolism in patients with obesity. This circulating exosomal miRNA signature in patients with obesity with or without DM is a potential biomarker and therapeutic target in these patients.
- obesity
- diabetes mellitus
- type 2
- biomarkers
- transcription
- genetic
This is an open access article distributed in accordance with the Creative Commons Attribution Non Commercial (CC BY-NC 4.0) license, which permits others to distribute, remix, adapt, build upon this work non-commercially, and license their derivative works on different terms, provided the original work is properly cited, appropriate credit is given, any changes made indicated, and the use is non-commercial. See: http://creativecommons.org/licenses/by-nc/4.0/.
Statistics from Altmetric.com
Significance of this study
What is already known about this subject?
MicroRNA (miRNA) changes in exsosome contribute to the development of diabetes or obesity.
What are the new findings?
We found the similarity in the exosomal miRNA profiles of patients with obesity with or without diabetes.
Our study demonstrated the specific circulating exosomal miRNAs of patients with obesity with diabetes compared with patients with obesity without diabetes and healthy volunteers.
Our study showed miRNAs that change sequentially in patients with obesity without diabetes and patients with obesity with diabetes compared with healthy volunteers.
How might these results change the focus of research or clinical practice?
This circulating exosomal miRNAs signature of patients with obesity and with or without diabetes is a potential biomarker and therapeutic target of obesity and obesity-related complications.
Introduction
Modern sedentary lifestyles and excessive calorie intake have increased the risk of developing metabolic diseases.1 Obesity is a major component of metabolic syndrome, which includes a cluster of metabolic disturbances such as hypertension, dyslipidemia, hyperglycemia, insulin resistance, obesity and inflammation.2 Obesity poses a substantial health risk, as it has been linked to several common diseases, such as type 2 diabetes mellitus (T2DM), cardiovascular disease, stroke, arthritis and several types of cancer.3 4 The upsurge in obesity rates appears to be the primary factor affecting the recent increase in the prevalence of T2DM.5 Although T2DM is associated with a strongly increased cardiovascular disease risk, the process of irreversible subclinical vascular damage begins in the prediabetes stage.6
T2DM creates a burden for patients and is a risk factor for many complications. Diabetes mellitus increases the risk of cardiovascular disease by threefold to fourfold in women and twofold to threefold in men after adjusting for other risk factors.7 There is a positive correlation between the volume of visceral fat tissue in patients with T2DM and arterial inflammation.8 Another study demonstrated that metabolic syndrome is associated with increased arterial stiffness in patients with T2DM, and abdominal obesity is associated increased arterial stiffness.9 Obesity and diabetes are risk factors for cardiovascular disease, and cardiovascular risk increases when both factors coexist.8–10 Therefore, preventing T2DM in obesity is critical issue for preventing cardiovascular complications. T2DM is a progressive disease with a course that typically depends on β-cell dysfunction and worsening of insulin resistance.11 However, the progression of diabetes from insulin resistance and the factors that play an important role in the process are not well-understood.
Recent studies have shown that exosomes are involved in multiple pathological processes, such as insulin resistance, lipid toxicity, dyslipidemia, endocrine disorders, hypercoagulable state and chronic inflammation.12–14 Most cells in the human body can secrete exosomes containing mRNA, microRNA (miRNA) and other noncoding RNAs14; exosomal miRNAs can be stabilized in circulation via protection from the exosome, and then are transferred into target cells to inhibit or activate the expression of certain genes.15 16 The mechanisms by which obesity causes diabetes and vascular complications have been described. miRNAs affect adipose tissue metabolism (adipogenesis, release and action of signals derived from adipose tissue) and insulin secretion and activity, and an imbalance in these miRNAs can lead to the development of obesity and related complications.17 Indeed, dysregulation of miRNAs is a common feature in various human diseases including atherosclerosis and obese/diabetes-associated vascular complications.18 The miRNAs carried by exosomes in the body fluids may act as efficient biomarkers for diabetic complications, such as diabetic nephropathy.14 19 Several recent studies have evaluated miRNAs in diabetes and the prediabetes state20; however, most studies focused on serum cell free circulating miRNAs using microarray or real-time quantitative PCR (qPCR) in prediabetic conditions and overt diabetes. Additionally, these studies did not focus on obesity.
Our previous work investigated the difference of circulating exosomal miRNA profiles between patients with obesity without T2DM and healthy volunteers (HVs).12 However, the effects of persistent high glucose such as in DM on circulating exosomal miRNAs in patients with obesity has not been evaluated. Therefore, we compared the miRNA levels through small RNA sequencing to identify differentially expressed miRNAs to highlight potential pathways involved in the progression of T2DM in patients with obesity, as well as to provide new targets for treating obesity-associated diabetes.
Research design and methods
Study participants
We prospectively recruited HVs and subjects with obesity at Soonchunhyang University Hospital (Seoul, South Korea). All participant’s demographic data and anthropometric parameters were obtained at the time of enrollment. Obesity was defined as body mass index (BMI) ≥30.0 kg/m2 according to the Steering Committee of the Regional Office for the Western Pacific Region of WHO. BMI was calculated by dividing the initial body weight (kg) by height (m) squared. DM was confirmed based on WHO criteria: fasting glucose ≥126 mg/dL, serum glucose ≥200 mg/dL after a 2-hour oral glucose tolerance test, hemoglobin A1c level ≥6.5%21 and diabetes medication history. Blood samples were obtained at least 8 hours after fasting, and the estimated glomerular filtration rate was calculated using the CKD Epidemiology Collaboration equation.22
Circulating exosomal RNA isolation and assessment
We isolated exosomes from serum using the ExoQuick isolation agent (System Biosciences, Palo Alto, California, USA), according to the manufacturer’s guidelines. Serum samples were centrifuged at 3000 ×g for 15 min; subsequently, supernatants were mixed with ExoQuick reagent and incubated for 30 min at 4°C. After centrifugation, the supernatants were aspirated. The remaining pellets contained exosomes. We resuspended the pellets in 100–200 µL of sterile phosphate-buffered saline and RNA was extracted with the miRNeasy Mini Kit (Qiagen, Hilden, Germany). Exosome suspensions (200 µL) were mixed with QIAzol lysis buffer (1 mL) (Qiagen), and the mixtures were processed according to the manufacturer’s guidelines. The RNA was eluted in RNase-free water (20 µL). Purified RNA was analyzed for exosomal RNA size distribution using an Agilent 2100 Bioanalyzer with an RNA Pico Chip and Small RNA Chip (Agilent Technologies, Santa Clara, California, USA).
Complementary DNA library preparation and small RNA sequencing
RNA sequencing was conducted as described previously.12 The samples were processed to produce exosomal RNA (10 ng) as input for each library. Small RNA libraries were constructed using the SMARTer smRNA-Seq Kit for Illumina (Takara Bio, Shiga, Japan) according to the manufacturer’s guidelines. We generated sequencing libraries by polyadenylation, complementary DNA synthesis and PCR amplification.
The libraries were gel-purified and validated based on their size, purity and concentration, using an Agilent Bioanalyzer. They were quantified by qPCR according to the qPCR Quantification Protocol Guide (KAPA Library Quantification Kits for Illumina Sequencing Platforms). We assessed the quality of libraries using a TapeStation D1000 ScreenTape (Agilent Technologies). Equimolar amounts of libraries were pooled and sequenced on an Illumina HiSeq 2500 instrument (Illumina, San Diego, California, USA) to generate 101 base reads. To perform image decomposition and quality value calculations, we used the modules in the Illumina pipeline. We conducted all procedures for next-generation sequencing analysis through Macrogen (Seoul, Korea).
Analysis of RNA sequencing data
Known and novel miRNAs were identified using the miRDeep2 software algorithm after sequence alignment. We retrieved the Homo sapiens reference genome release hg19 from the UCSC Genome Browser, which was indexed using Bowtie (1.1.2), a program for aligning experimental and reference sequences before sequencing alignment. The reads were then aligned to the mature and precursor H. sapiens miRNAs, obtained from miRBase 21.
Proportion of miRNAs and other RNAs
Uniquely clustered reads were sequentially aligned to the reference genome using miRBase 21 and non-coding RNA database Rfam 9.1 to identify known miRNAs and other types of RNAs, respectively.
Analysis of miRNA expression levels
The raw data (reads for each miRNA) were normalized by the relative log expression using DESeq2. For preprocessing, miRNAs absent from >50% of the samples were excluded, leaving mature miRNAs for analysis. We added one to the normalized read count of the filtered miRNAs and facilitated log2 transformation in order to draw a correlation plot. For each miRNA, the baseMean and log fold-change were calculated between case and control. For comparison of the two groups, we conducted a statistical hypothesis test by using the negative binomial Wald test in DESeq2, and defined the miRNAs that differentially expressed between the two groups as having a |fold change|≥2 and false discovery rate-adjusted p value <0.05. We also performed a hierarchical clustering analysis, using complete linkage and Euclidean distance as measures of similarity, to display the expression patterns of differentially expressed miRNAs that satisfied the criteria of fold-change|≥2 and false discovery rate-adjusted p value of<0.05. All data analyses and visualization of differentially expressed genes were conducted using R V.3.3.1 (www.r-project.org).
miRNA target prediction and functional annotation analysis
To evaluate the functions of the differentially expressed miRNAs, miRNA target prediction was performed using the miRSystem database. The miRSystem used seven algorithms for predicting miRNA targets (namely, DIANA-microT, miRanda, miRBridge, PicTar, PITA, RNA22 and TargetScan) and two experimentally validated databases of miRNA target genes (TarBase and miRecords). The gene list of predicted targets (prediabetes only or diabetes only miRNA) submitted to the Database for Annotation, Visualization, and Integrated Discovery and the putative targets were annotated using four pathway databases, including Kyoto Encyclopedia of Genes and Genomes, Biocarta, Pathway Interaction Database and Reactome.
Statistical analyses
Continuous variables are expressed as the mean±SD, and categorical variables are expressed as a number and percentage. Variables without normal distribution are expressed as the median and IQR. Continuous variables of HVs and patients with obesity were analyzed by the Mann-Whitney U test. Categorical and non-parametric variables were compared using a χ2 test and Kruskal-Wallis test, as appropriate. Statistical analyses were performed using SPSS V.23.0 (IBM, Armonk, New York, USA).
Results
Baseline characteristics
A total of 47 participants (18 HVs, 16 patients with obesity without DM, and 13 patients with obesity with DM), who voluntarily provided informed consent, were enrolled. Baseline characteristics of the participants are shown in table 1. All measurements except diastolic blood pressure, blood urea nitrogen, creatinine, urinary protein and urinary albumin differed between HVs and patients with obesity without DM. The patients with obesity with DM and the HVs were older compared with the patients with obesity without DM. Gender, blood pressure, weight, BMI, waist:hip ratio, serum creatinine, cystatin C, lipid profiles and C-peptide did not differ between the groups with obesity. No participant had hypertension, with the exception of 10 patients (76.9%) with DM and obesity. No other participants had cerebrovascular disease or cancer.
Baseline characteristics of study participants
Profiles of exosomal miRNAs among participants
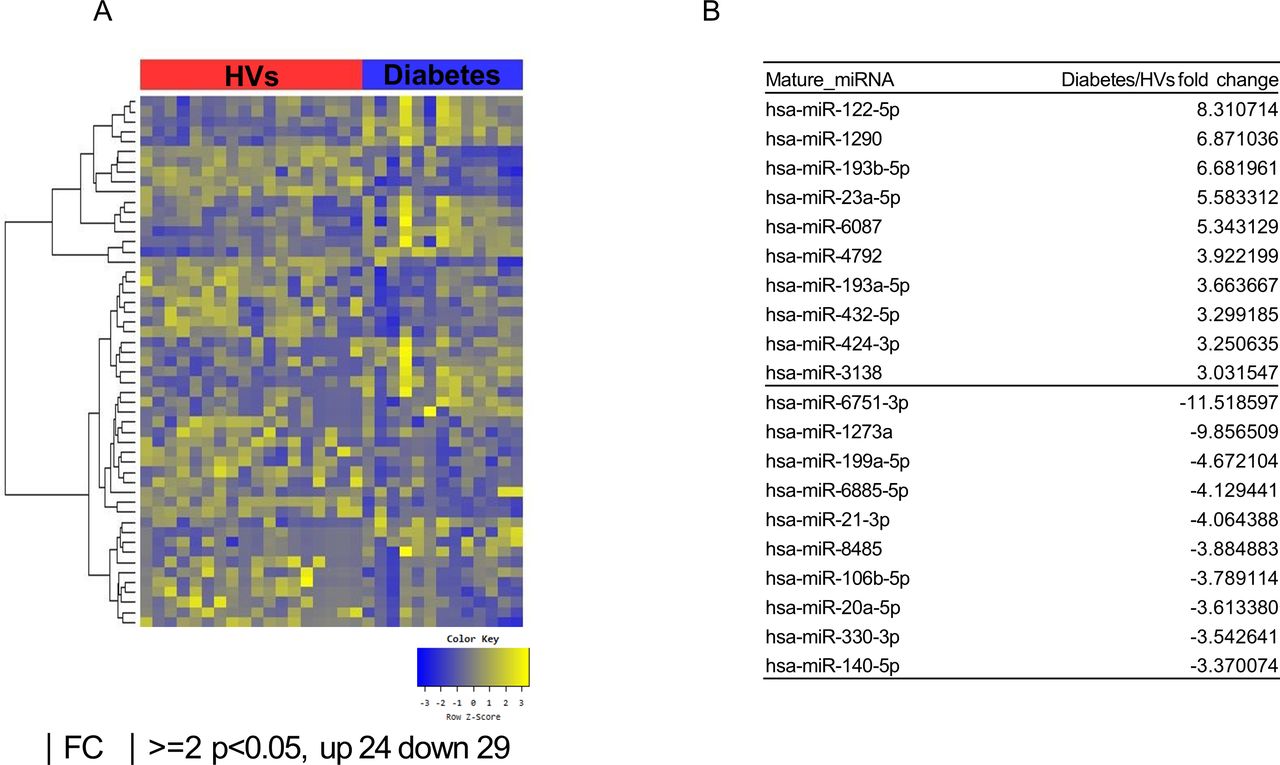
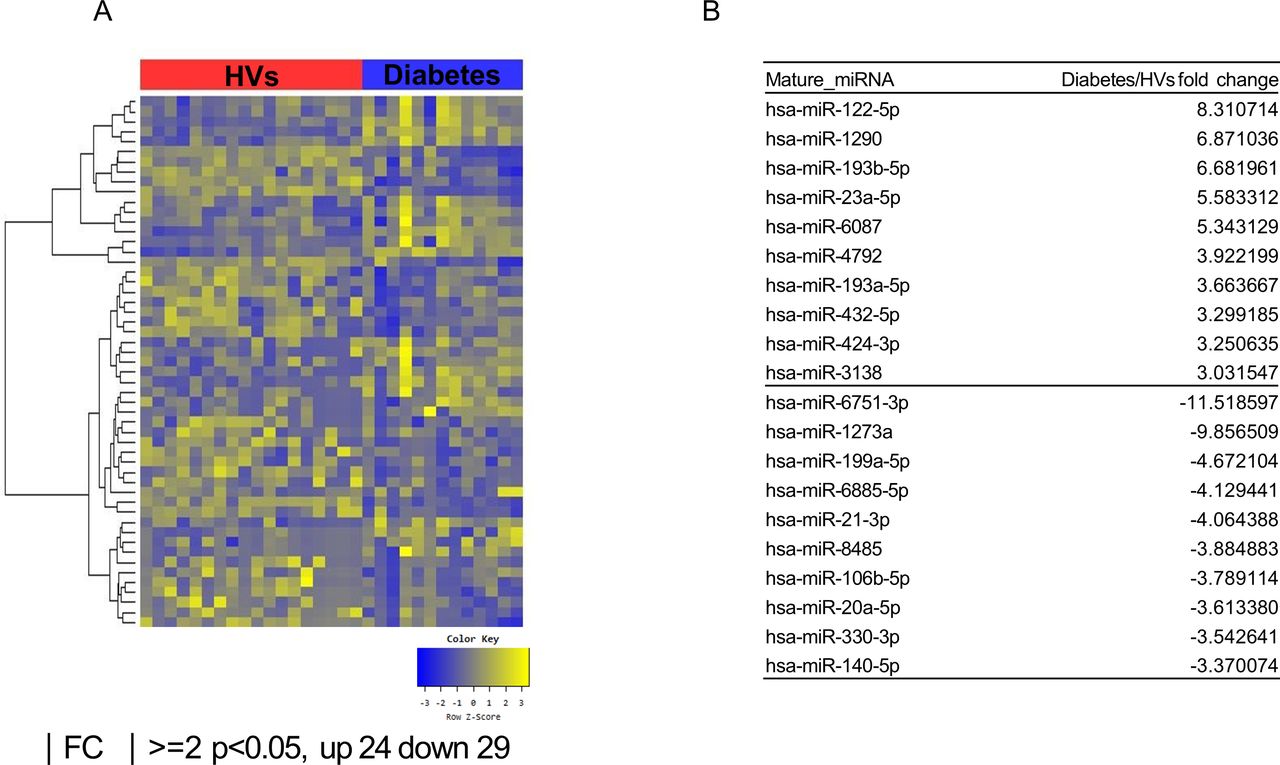
According to the multidimensional scaling graph, the patients with obesity with DM and patients with obesity without DM had many overlapping areas (online supplementary figure S1). After small RNA sequencing, we identified 26 upregulated and 28 downregulated miRNAs in patients with obesity without DM compared with HVs (figure 1A,B). We also found 24 upregulated and 29 downregulated miRNAs in patients with obesity with DM compared with those of HVs (figure 2A,B). Comparison of the patients with obesity without DM and patients with obesity with DM revealed 14 upregulated miRNAs and 11 downregulated miRNAs in patients with obesity with DM (figure 3A,B). To identify unique miRNAs in patients with obesity without DM or patients with obesity with DM, we selected miRNAs upregulated only in one group compared with HVs (table 2 and online supplementary figures S2 and S3).
Supplemental material
Supplemental material
Supplemental material
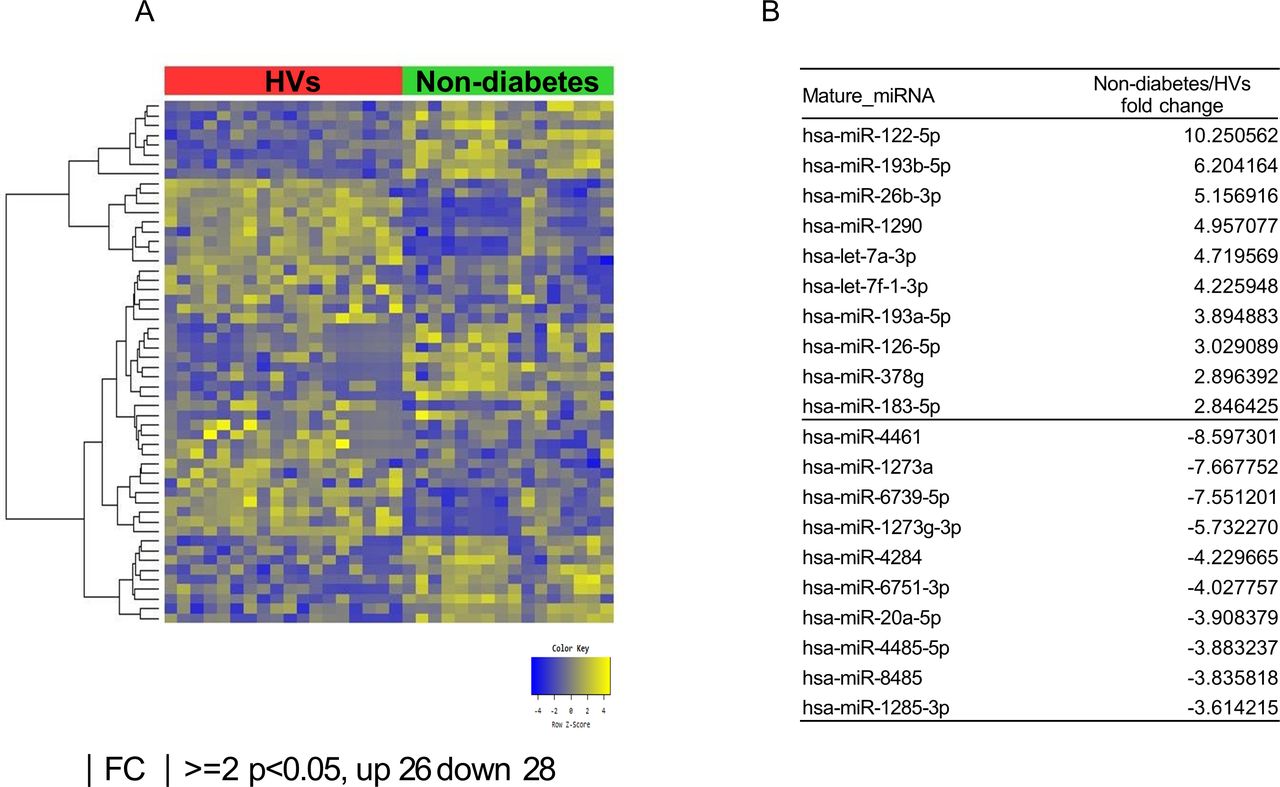
Exosomal microRNAs (miRNAs) from patients with obesity without DM differ from those from healthy volunteers (HVs). (A) Heat map showing z-score of exosomal miRNAs from HVs (n=18) and patients with obesity without DM (n=16) with 26 upregulated (yellow) and 28 downregulated (blue) miRNAs. (B) Fold change (FC) of top 10 miRNA expression in patients with obesity without DM compared with HVs. DM, diabetes mellitus.
Exosomal microRNAs (miRNAs) from patients with obesity with DM differ from those from patients with obesity without DM. (A) Heat map showing z-score of exosomal miRNAs from patients with obesity without DM (n=16) and patients with obesity with DM (n=13) with 14 upregulated (yellow) and 11 downregulated (blue) miRNAs. (B) Fold change (FC) of top 10 miRNA expression in patients with obesity with DM compared with patients with obesity without DM. DM, diabetes mellitus.

{kind=link}
{kind=link}
{kind=link}
Exosomal microRNAs (miRNAs) from patients with obesity with DM differ from those from healthy volunteers (HVs). (A) Heat map showing z-score of exosomal miRNAs from HVs (n=18) and patients with obesity with DM (n=13) with 24 upregulated (yellow) and 29 downregulated (blue) miRNAs. (B) Fold change (FC) of top 10 miRNA expression in patients with obesity with DM compared with HVs. DM, diabetes mellitus.
Upregulated exosomal miRNAs only in patients without diabetes/patients with diabetes compared with HVs
Exosomal miRNAs associated with T2DM development in obesity
We identified the specific miRNAs that continuously changed from HVs to patients with obesity without and with diabetes. Expression of miR-23a-5p in patients with obesity without DM increased by 2.12-fold compared with HVs; the value in the patients with obesity with DM was 2.64-fold higher than that in patients with obesity without DM. Additionally, miR-6087 levels in patients with obesity without DM were 2.64-fold higher than those in HVs, whereas these values in patients with obesity with DM increased 2.03-fold compared with those patients with obesity without DM. In contrast, miR-6751-3p levels in patients with obesity without DM were 4.03-fold lower than those in HVs, whereas these values in patients with obesity with DM decreased 2.86-fold compared with those in patients with obesity without DM.
Prediction of targets and functional annotation of obesity with or without diabetes-specific miRNAs
We identified 15 obesity-specific miRNAs and 9 diabetes-specific miRNAs with significantly different expression compared with HVs (table 2). Targets of these core miRNAs were predicted using miRSystem. Obesity-specific miRNAs and diabetes-specific miRNAs were predicted to target 1543 and 2405 genes, respectively. The possible pathways targeted by these miRNAs are presented in online supplementary figures S4 and S5.
Supplemental material
Supplemental material
Discussion
In this study, we found that the miRNA profiles in exosomes of patients with obesity with DM were similar to those in patients with obesity without DM compared with HVs. These results suggest that obesity has an overwhelming effect on exosomal miRNAs and that these changes compared with individuals without obesity contribute to metabolic complications. We also identified the miRNAs that differed according to presence of diabetes, which may play an important role in the pathological mechanism by which patients with insulin resistance develop diabetes.
Recent in vivo studies demonstrated that exosomes can transfer mature miRNAs between organs, resulting in functional changes in the receiving cells and affecting whole-body insulin sensitivity.13 23 The transfer of obesity-associated exosomal miRNAs induced glucose intolerance, adipose inflammation and hepatic steatosis in lean mice.13 These results support a role for obesity-related exosomal miRNAs in metabolic complications. The similarity of exosomal miRNAs in the patients with obesity without DM and patients with obesity with DM in our study also provide insight into the pathogenesis of vascular complications in obesity. miR-122–5 p, miR-193b-5P, miR-193a-5P and miR-1290 were the most upregulated miRNAs in the patients with obesity compared with HVs, whereas miR-1273a and miR-20a-5p were the most downregulated miRNAs. These miRNAs may contribute to the pathogenesis of vascular complications in the patients with obesity and to insulin resistance, thus contributing to the development of diabetes. For example, miR-122–5 p is known to be associated with the prognosis of acute myocardial infarction24 and is a fatty liver and cardiovascular risk factor.25 One study reported that miR-20a-5p is significantly decreased in gestational diabetes and is associated with glucose homeostasis, including insulin signaling, phosphatidyl-inositol-3-kinase (PI3K)-protein kinase B (Akt), and forkhead box protein O (FoxO).26 Both of miR-193a-5P and miR-193b-5P have been reported to contribute to tumor suppressor in various cancers.27 miR-1290 has also been studied mainly in cancer, and most studies showed that overexpression of miR-1290 increases cancer cell proliferation and invasion.28 Reduction of miR-1273a is also known to increase the activity of circular PIP5K1A associated with progression of colon cancer.29 These results support that obesity is associated with cancer development via miRNAs and that these miRNAs should be validated in a cancer cohort.
We determined the exosomal levels of 15 obesity-specific miRNAs. One-third of these miRNAs belongs to the let-7 family (let-7b-3p, let-7f-1-3p, let-7d-3p, let-7a-3p and let-7c-5p), which is downregulated in the development of diabetes and vascular complications.30 Although miR-126-5p is known to have a protective effect on atherosclerosis, miR-192-5p has been shown to protect the kidney from damage caused by hypertension.31 32 Urinary exosomal miR-21-5p is increased in diabetic nephropathy, and is associated with kidney damage caused by diabetes.33 In addition, miR-29b-3p was found to be specific to islet beta-cell stress.34 This is the first study to report that the other six miRNAs (miR-378g, miR-556-3p, miR-151a-3p, miR-130b-5p, miR-4488 and miR-183-5p) are associated with metabolic syndrome or diabetes. The changes in these miRNAs indicate that vascular complications are already occurring in patients with obesity without DM. Further studies are needed to determine whether the newly discovered miRNAs play a role in the promotion or inhibition of the progression of metabolic syndrome to diabetes.
Additionally, there were nine diabetes-specific miRNAs, which are thought to play a key role in the development of diabetes and vascular complication. Some miRNAs were reported to be associated with hyperglycemia or vascular disease. In a study comparing patients with T2DM with healthy individuals, miR-3960 was one of five miRNAs that was significantly different in serum.35 One study showed that N-acetyl-seryl-aspartyl-lysyl-proline (AcSDKP) associated with the fibroblast growth factor receptor 1 signaling pathway maintained endothelial mitochondrial homeostasis by inducing miR-let-7b-5p expression.36 Additionally, overexpression of miR-320a is a key risk factor for atherosclerosis37 and another study showed that overexpression of miR-320b abolishes enhanced mesenchymal stem cell-medicated cardioprotection.38 Five other diabetes-specific miRNAs have not been reported to be directly related to obesity or diabetes.
Two miRNAs were observed to be sequentially increased in patients with obesity without DM and patients with obesity with DM compared with HVs; the only sequentially reduced miRNA was considered responsible for the mechanism of diabetes development. One of these miRNAs, miR-23a-5p, has been found to be associated with atherosclerosis plaque progression and vulnerability by promoting foam cell formation. This miRNA attenuates cholesterol efflux to promote foam cell formation by downregulating the expression of ATP-binding cassette transporters (ABCA1 and ABCG1).39 The other, miR-6087, is known to inhibit endoglin expression, a type of transforming growth factor-β receptor on endothelial cells. It is known to play an important role in angiogenesis,40 and is associated with expression of the tumor suppressor gene p53.41 Recent studies showed that p53 gene polymorphisms affect life expectancy and lifestyle-related diseases, such as T2DM and obesity, indicating a relationship between p53 function and metabolic disorders.42 No studies have examined the correlation between miR-6751-3p and diabetes, and the function of miR-6751-3p is unknown. Diabetes does not develop in all patients with obesity, but the continuously changing miRNAs identified in our study appear to play an important role in diabetes development in patients with obesity. Further studies are needed to clarify the functions of miRNA in diabetes.
Pathway analysis related to miRNAs revealed common biological pathways in both the patients with obesity with DM and patients with obesity without DM. As expected, diabetes-specific miRNAs significantly affect the pathways associated with insulin secretion, insulin regulation and energy metabolism. In the patients with obesity without DM, miRNAs significantly affected pathways associated with insulin secretion, inflammatory cytokine, interferon, muscle atrophy and vascular smooth muscle atrophy. The insulin receptor, WNT, AMP-activated protein kinase, transforming growth factor-β signaling and mitogen-activated protein kinase (MAPK) signaling pathways were among the top canonical pathways, identified in previous studies of metabolic syndrome.12 43 Pathway analysis in our current study was similar to that in our previous study.12 19 Representative examples of important pathways in our study include the MAPK signaling and integrin pathways. MAPK signaling pathway, axon guidance signaling pathway, neurotrophin signaling pathway and notch signaling pathway are associated with insulin resistance.44 MAPK signaling is known to play an important role in diabetic nephropathy.19 In addition, the integrin-vascular endothelial growth factor axis is a key factor in pathological angiogenesis, including in tumor metastasis, tissue remodeling and diabetic complications.19 Further studies are required to evaluate the association between these miRNAs and the biological pathways identified, as well as the mechanisms and critical points in the development of diabetes from metabolic syndrome.
There were some limitations to our study. First, a relatively small number of participants was enrolled in this study. Further studies with a larger cohort are warranted to confirm our findings. Second, some demographic factors and clinical parameters differed between groups, which may have affected the circulating exosomal miRNA profiles. Third, although we conducted exosomal RNA sequencing using validated commercial kits, the procedures used to isolate the exosomes are not standardized, and systematic bias in library preparation may lead to false-positive identification of miRNAs.
Conclusions
We identified specific miRNAs and associated biological pathways in patients with obesity without DM and patients with obesity with DM. Moreover, specific exosomal miRNAs of patients with obesity without DM and patients with obesity with DM may provide clues to the mechanism of hyperglycemic complications. Adipokines, inflammation and reactive oxygen species in patients with obesity may affect the cells in the body, resulting in exosomal miRNA changes, leading to developing diabetes and vascular complications; however, further research is needed to confirm this. In conclusion, these miRNAs can be considered as candidates for treating metabolic disease and diabetes.
References
Footnotes
HyosK and Y-UB contributed equally.
Contributors HyosK contributed to data analysis, statistical analysis, literature search and writing the manuscript. Y-UB involved in data analysis, conduction of the experiments, statistical analysis and cowriting the manuscript. HL, HyouK, JSJ, HN, DCH and DWB involved in data analysis, searching literature and editing the manuscript. SaHK and HKP involved in study design, data collection and editing the manuscript. SR and SoHK contributed to study design, submitting the grant, conduction of the experiments, data collection and analysis, literature search and editing the manuscript.
Funding This study was supported by the Bio & Medical Technology Development Program of the National Research Foundation (NRF) funded by the Korean government (MSIT) (NRF-2019M3E5D3073092) and the Soonchunhyang University Research Fund (20191124).
Competing interests None declared.
Patient consent for publication Not required.
Ethics approval This study was conducted in accordance with the Declaration of Helsinki, and the study protocol was approved by the institutional review board of Soonchunhyang University Seoul Hospital (IRB No: 2015-11-020).
Provenance and peer review Not commissioned; externally peer reviewed.
Data availability statement Data are available on reasonable request.