Article Text

Abstract
Introduction We previously reported in ob/ob mice, one of animal models of human type 2 diabetes mellitus (DM2), that (i) acetylation of histone H3 lysine 9 (H3K9) at the promoter region of clock gene Dbp and DBP mRNA expression are reduced in epididymal adipose tissue, (ii) binding of DBP to the promoter region of peroxisome proliferator-activated receptor (Ppar)-γ and mRNA expression of PPAR-γ1sv were decreased in preadipocytes and (iii) adiponectin secretion was decreased, leading to the impaired insulin sensitivity.
Research design and methods The present study was undertaken to evaluate whether such the changes in visceral adipose tissue were detected in patients with DM2. We obtained omental and mesenteric adipose tissue during surgery of lymph node dissection for gastric and colorectal cancers, and investigated these variables in adipose tissue (omental from gastric cancer; 13 non-DM, 12 DM2: mesenteric from colorectal cancer; 12 non-DM, 11 DM2).
Results Acetylation of histone H3K9 at the promoter region of Dbp and DBP mRNA expression in omental, but not in mesenteric adipose tissue were significantly lower in DM2 than in patients without DM. PPAR-γ mRNA expression in omental adipose tissue was also lower in patients with DM2, but not in mesenteric adipose tissue.
Conclusions The changes in DBP-PPAR-γ axis observed in mice with diabetes were also detected in patients with DM2. Because adiponectin secretion is reported to be enhanced through the PPAR-γ-related mechanism, this study supports the hypothesis that omental adipose tissue is involved in the mechanism of DM2.
- gene expression
- circadian rhythm
This is an open access article distributed in accordance with the Creative Commons Attribution Non Commercial (CC BY-NC 4.0) license, which permits others to distribute, remix, adapt, build upon this work non-commercially, and license their derivative works on different terms, provided the original work is properly cited, appropriate credit is given, any changes made indicated, and the use is non-commercial. See: http://creativecommons.org/licenses/by-nc/4.0/.
Statistics from Altmetric.com
Significance of this study
What is already known about this subject?
Disruption of circadian rhythms is associated with the increased risk of type 2 diabetes mellitus (DM2).
Histone acetylation level at the promoter regions of Dbp gene, a molecular clock component and DBP mRNA expression are significantly reduced in epididymal adipose tissue of mice with obesity having diabetes.
What are the new findings?
Histone acetylation at the promoter regions of Dbp in omental adipose tissue was significantly reduced in patients with DM2.
In omental adipose tissue, mRNA expressions of DBP and novel peroxisome proliferator-activated receptor-γ were significantly lower in patients with DM.
These changes were not detected in mesenteric adipose tissue.
How might these results change the focus of research or clinical practice?
Omental adipose tissue is involved in the pathophysiology of DM2, rather than mesenteric adipose tissue.
Introduction
Circadian clock regulates 24 hours rhythms which are essential to maintaining normal human physiology including metabolic functions. Disruption of circadian rhythms (eg, shift work and sleep disorders) is associated with the increased risk of type 2 diabetes mellitus (DM2) and obesity.1 2
Circadian rhythms are controlled by a molecular clock system located in the hypothalamic suprachiasmatic nucleus (center clock), and in the peripheral tissue (peripheral clock).3 The molecular clock system is composed of genes such as Clock, brain and muscle ARNT-like 1 (Bmal1), period (Per) and cryptochrome (Cry), which constitute an oscillatory mechanism based on self-sustaining transcriptional/translational feedback loops.3–6 Comprehensive network circuits are also regulated through the actions of E-box/E’-box, DBP/E4BP4 binding element (D-site) and Rev-erbα/ROR binding element.7
Albumin D-site-binding protein, known as DBP, is a molecular clock component.8 We have previously reported that the acetylation of histone H3 lysine 9 (H3K9) at the promoter regions of Dbp gene and DBP mRNA expression in mice with obesity having diabetes (ob/ob) are significantly reduced in epididymal adipose tissue.9 Recently, we showed that the binding of DBP to the promoter region of novel peroxisome proliferator-activated receptor (Ppar)-γ and the mRNA expression of PPAR-γ1sv, a splicing variant of PPAR-γ, were decreased in the fractionated preadipocytes of ob/ob mice.10 It subsequently decreased the secretion of adiponectin, leading to the impaired insulin sensitivity in ob/ob mice. Because ob/ob mice is one of animal models of human DM2,11 we hypothesized that the acetylation of histone H3K9 at the promoter region of Dbp gene is blunted, and subsequent mRNA expressions of DBP and human novel PPAR-γ isoform, which is homolog corresponding to mouse PPAR-γ1sv,12 are decreased in visceral adipose tissue in patients with DM2. This study was undertaken to address the issues in patients with DM2.
Omental and mesenteric adipose tissue are two major components of visceral fat. Omentum is the fatty apron attached to the greater curvature of the stomach, and mesentery is the fat containing the arteries and veins supplying the small and large intestine.13 Because physiological and pathological profiles are reported to differ between omental and mesenteric adipocytes in patients with obesity,13 14 it remains probable that the profiles of DBP differ between omental and mesenteric adipose tissue in patients with DM2. To evaluate the hypothesis, DBP mRNA expressions in two different adipose tissues were also compared in this study.
Materials and methods
Patients
Patients with gastric and colorectal cancers were enrolled in this study. They underwent lymph node dissection for gastric (n=26) and colorectal (n=24) cancers in the Division of Gastroenterological Surgery, Jichi Medical University. On the day before surgery, all patients admitted to the hospital before noon and took their lunch. Thereafter, they did not eat any foods before surgery. Light-on time is 06:00 hours and light-off time is 21:00 hours in a hospital room. Patients who were scheduled to begin their operation at 09:00 were enrolled to ensure a collection of all adipose tissue within a short time window. The exclusion criteria were as follows: (i) age <20 years, (ii) patients who could not provide their informed consent and (iii) patients with corticosteroid or metformin medication, which are known to influence clock gene expression.15 16
Number of patients enrolled in every 3 months are shown in online supplementary table 3. There were no significant seasonal differences between the patients without DM2 and patients with DM2 in the gastric or colorectal cancer group. Patients characteristics are shown in table 1. The values of hemoglobin A1c (HbA1c) and/or fasting blood glucose were higher in the DM2 group than in the non-DM group in patients with gastric and colorectal cancer. There were no significant differences in other parameters between the DM2 and non-DM groups in patients with gastric and colorectal cancer. Oral antidiabetic drugs were stopped on the day of surgery.
Supplemental material
Patient characteristics
Sample collection
The removal of organs including the adipose tissue of omental (in patients with gastric cancer) and mesenteric (in patients with colorectal cancer) was performed by a gastrointestinal surgeon in the operating room. Thereafter, pathologists (HKa or AT) immediately dissected the omental and mesenteric adipose tissue for RNA extraction and chromatin immunoprecipitation (ChIP) analysis.
The median o’clock time of omental and mesenteric adipose tissue dissection was 12:10 and 12:30, respectively. Almost all sampling was finished within 90 min after the initiation of surgery (10:40–13:40). Because samples of one patient without diabetes with gastric cancer and one patient without diabetes with colorectal cancer were collected at 14:00 and 15:00, respectively, data of these patients were not included for all analyses. In addition, because sufficient chromatin was not obtained from one patient with DM2 and three patients without diabetes with gastric cancer, these samples were not assessed via ChIP analysis.
RNA extraction and real-time PCR
Total RNA from adipose tissue were extracted using the RNeasy mini kit (Qiagen, Valencia, California, USA). Reverse transcription was performed using the QuantiTect Reverse Transcription kit (Qiagen). Real-time PCR was performed using the SYBR Select Master Mix (Thermo Fisher Scientific, Carlsbad, California, USA) and the Stratagene Mx3005P QPCR System (Agilent Technologies, Santa Clara, California, USA). Specific gene primer sequences are described in online supplementary table 1. Expression value of mRNA expression was normalized to the expression of ribosomal protein S18, and quantified using comparative threshold cycle method.
Chromatin immunoprecipitation
Adipose tissue was fixed with 1% formaldehyde for 15 min at room temperature. Fixed samples were homogenized and cross-linked chromatin was sonicated using a Bioruptor (Cosmo Bio, Tokyo, Japan) on ice. ChIP was performed using Dynabeads Protein G (Thermo Fisher Scientific). Fragmented chromatin was incubated overnight at 4°C with an antibody against acetyl-histone H3K9 (Cell Signaling Technology, Danvers, Massachusetts, USA). Antibody-reacted chromatin was purified using proteinase K and phenol-chloroform extraction. Purified DNA was subjected to real-time PCR amplification using SYBR Select Master Mix. Specific primers described in online supplementary table 2 were designed as follows: surrounding E-boxes in the promoter region of Dbp gene (H3K9 acetylation). Non-specific amplification was determined using an antibody against rabbit-IgG (Cell Signaling Technology).
Statistical analysis
The student’s t-test, Fisher’s exact test or Mann-Whitney U test were used to compare the groups. Correlations between the groups were analyzed by Spearman’s rank analysis. All calculations were performed using GraphPad Prism 6 (GraphPad Software, La Jolla, California, USA). Data were shown as median with IQR. The p value <0.05 was considered statistically significant.
Results
Clock genes mRNA expressions in omental and mesenteric adipose tissue
DBP mRNA expression was significantly (p<0.01) lower in the DM2 group than in the non-DM group in omental adipose tissue (figure 1A). However, an equivalent difference was not detected in mesenteric adipose tissue (figure 1B). The mRNA expressions of other clock genes measured in the study did not significantly differ between the DM2 and non-DM groups in visceral adipose tissue (figure 1).

Clock genes mRNA expressions in omental (A) and mesenteric (B) adipose tissue in the type 2 diabetes mellitus (DM2) and non-DM groups. Median with IQR, omental; n=13 in non-DM and n=12 in DM2, mesenteric; n=12 in non-DM and n=11 in DM2.
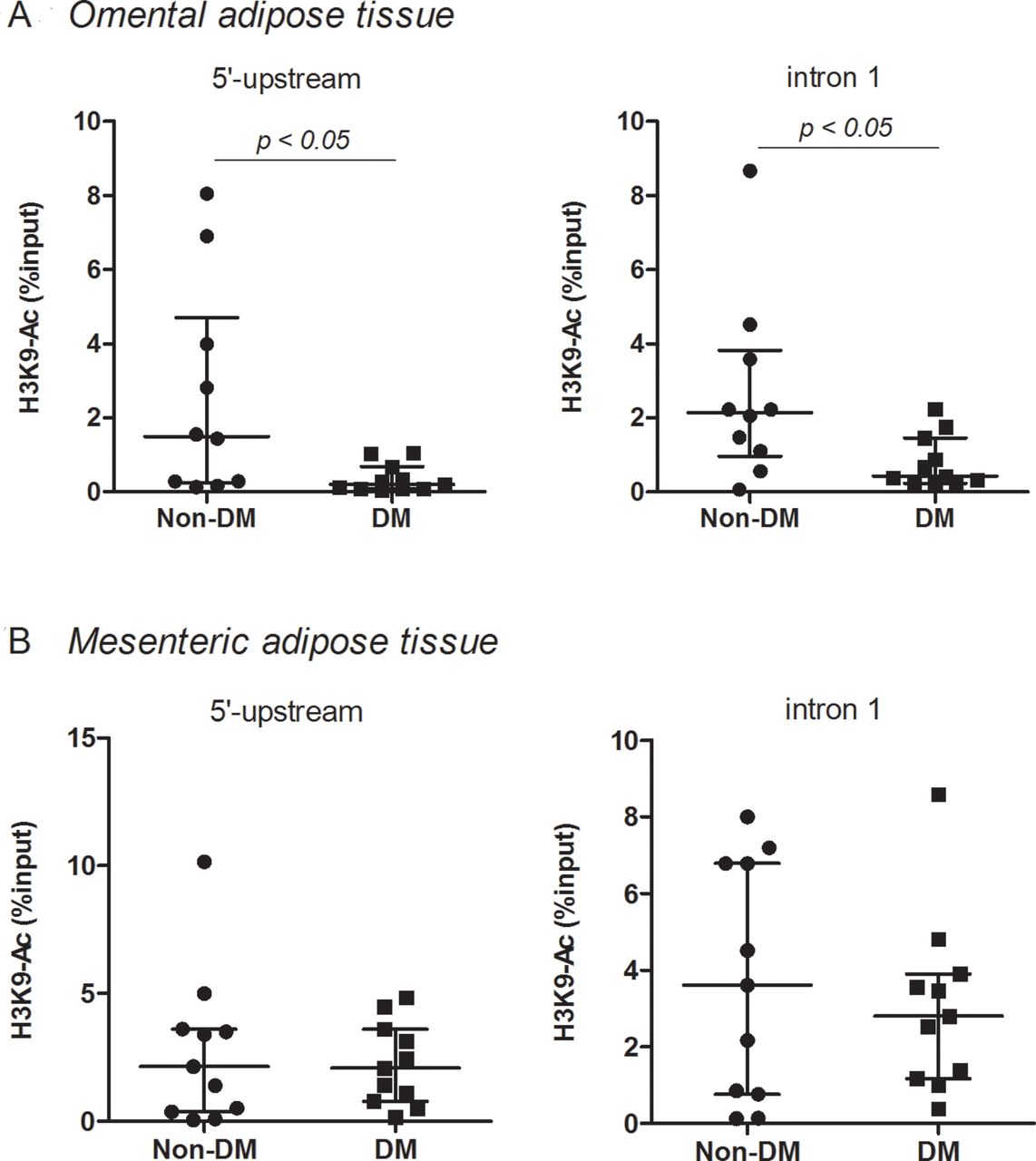
H3K9 acetylation at the promoter region of Dbp in omental and mesenteric adipose tissue
H3K9 acetylation at the promoter region (5’-upstream and intron 1) of Dbp in omental adipose tissue was significantly (p<0.05) reduced in the DM2 group compared with in the non-DM group (figure 2A). However, in mesenteric adipose tissue, H3K9 acetylation at the promoter region of Dbp in the DM2 group was not different from that in the non-DM group (figure 2B).

H3 lysine 9 (H3K9) acetylation at the promoter region of Dbp in omental (A) and mesenteric (B) adipose tissue in the type 2 diabetes mellitus (DM2) and non-DM groups. Median with IQR, omental; n=10 in non-DM and n=11 in DM2, mesenteric; n=12 in non-DM and n=11 in DM2.
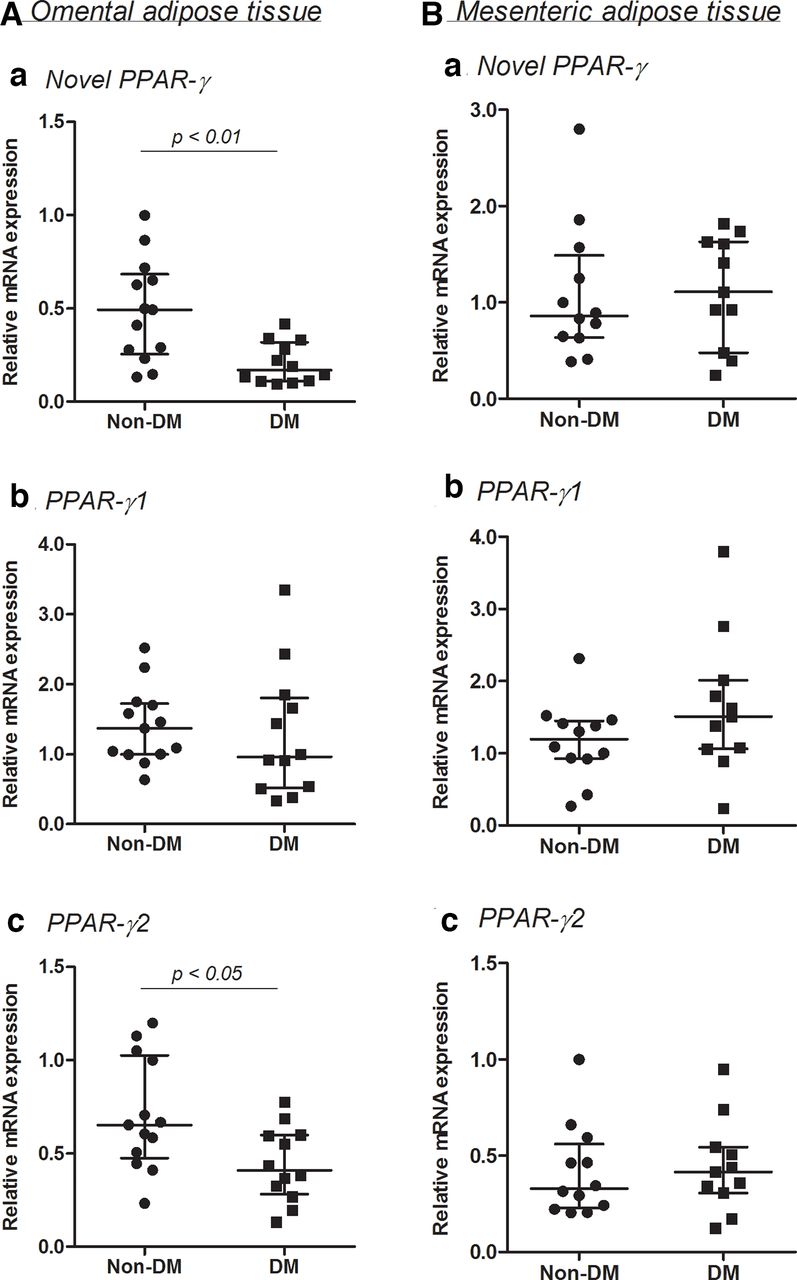
The mRNA expressions of three PPAR-γ isoforms in omental and mesenteric adipose tissue
The mRNA expression of PPAR-γ2 was higher than that of novel PPAR-γ and PPAR-γ1 in omental and mesenteric adipose tissue. The mRNA expressions of novel PPAR-γ and PPAR-γ1 were similar in these adipose tissue.
The novel PPAR-γ mRNA expression in the DM2 group was significantly (p<0.01) lower than that in the non-DM group in omental (figure 3A) but not in mesenteric adipose tissue (figure 3B). PPAR-γ2 mRNA expression in the DM2 group was also significantly (p<0.05) lower in omental, but not in mesenteric adipose tissue. PPAR-γ1 mRNA expressions of the DM2 and non-DM groups did not significantly differ in visceral adipose tissue (figure 3).

The mRNA expressions in three peroxisome proliferator-activated receptor (PPAR)-γ isoforms in omental (A) and mesenteric (B) adipose tissue in the type 2 diabetes mellitus (DM2) and non-DM groups. Median with IQR, omental; n=13 in non-DM and n=12 in DM2, mesenteric; n=12 in non-DM and n=11 in DM2.
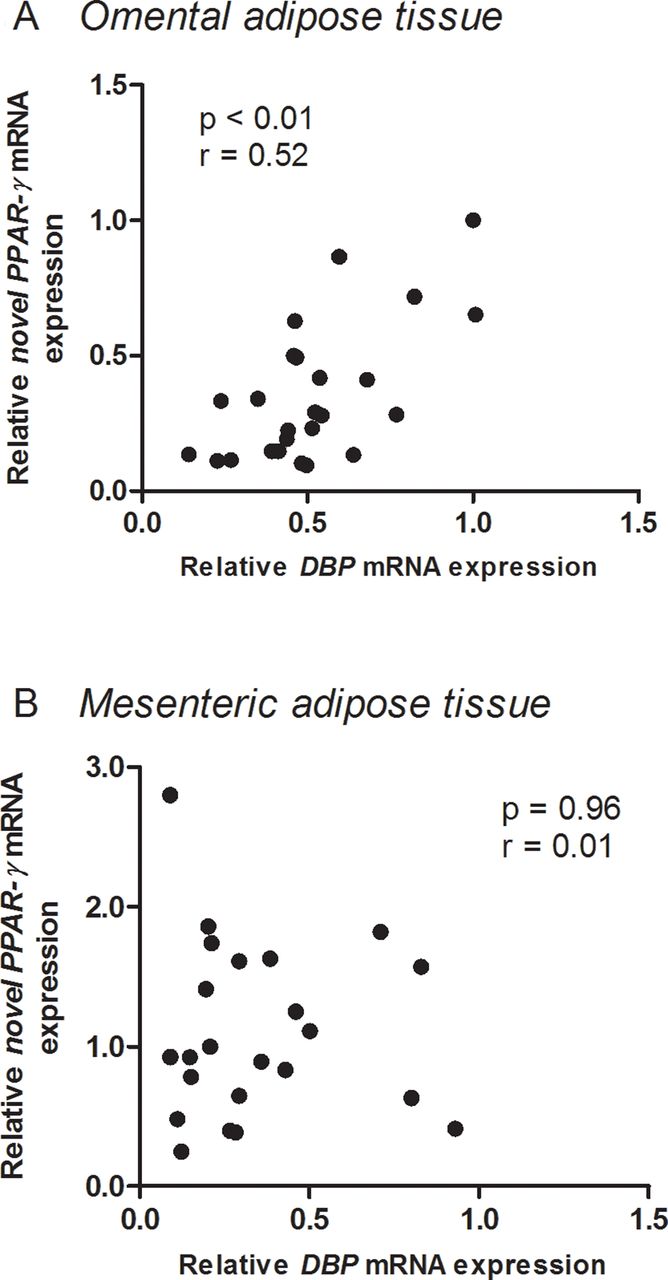
Correlation between DBP mRNA and novel PPAR-γ mRNA expressions in omental and mesenteric adipose tissue
There was a significant (p<0.01) positive correlation between DBP mRNA and novel PPAR-γ mRNA expressions in omental (figure 4A), but not in mesenteric (figure 4B) adipose tissue.
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Correlations between DBP mRNA and novel peroxisome proliferator-activated receptor (PPAR)-γ mRNA expressions in omental (A) and mesenteric (B) adipose tissue.
Discussion
The following data were obtained in omental adipose tissue in this study: (i) H3K9 acetylation at the promoter regions of Dbp was significantly reduced in the DM2 group compared with in the non-DM group; (ii) the mRNA expressions of DBP and novel PPAR-γ were significantly lower in the DM2 group than in the non-DM group and (iii) positive correlation was detected between DBP mRNA and novel PPAR-γ mRNA expressions.
It is well known that histone modifications, such as acetylation, exert a major role of transcriptional activity of downstream genes,17 and that a high intensity of H3K9 acetylation is detected around the transcription start site of downstream genes.18 19 Our previous studies showed that acetylation level of histone H3K9 at the promoter region of Dbp and DBP mRNA expression in epididymal adipose tissue were significantly reduced in ob/ob mice, one of the animal models of human DM2.9 10 We also found that histone deacetylases (HDACs) inhibitors increased DBP mRNA expression and decreased blood glucose in these animals with diabetes. In healthy cells, there is a tightly controlled equilibrium between the effects of histone acetyltransferases (HATs) and HDACs enabling histone (de)acetylation and the dynamic control of gene transcription.20 21 In this study, we demonstrated for the first time that H3K9 acetylation at the promoter regions of Dbp in omental adipose tissue was significantly reduced in patients with DM2. However, a mechanism of the reduction of H3K9 acetylation is not clear and remains to be determined. An imbalance between the activities of HATs and HDACs in favor of histone deacetylation is considered to be pathogenic in disease progression,20 21 which leads us to speculate that an imbalance between the activities of HATs and HDACs in the promoter regions of Dbp is also involved in the etiology of DM2. To address the issue, further clinical studies using drugs such as HDACs inhibitor are warranted in patients with DM2.
Tissue distribution and functions of PPAR-γ isoforms in mice are as follows: PPAR-γ1 is ubiquitously expressed except for adipose tissue and is involved in organ development such as placenta and heart. PPAR-γ1sv, a homolog corresponding to human novel PPAR-γ, and PPAR-γ2 are adipocyte-specific and synergistically regulate the early stage of adipocyte differentiation.12 22 In this study, only novel PPAR-γ has altered expression in omental adipose tissue in patients with DM2 with gastric cancer. However, there were no data which might explain the finding.
In this study, mRNA expressions of DBP and novel PPAR-γ were significantly lower in the DM2 group, which are similar with previous findings obtained in ob/ob mice.10 DBP is reported to bind directly to D-sites of promoter region of Ppar-γ and enhance the mRNA expression of the gene.23 Because the significant positive correlation was detected between DBP mRNA and novel PPAR-γ mRNA expressions, the reduction in novel PPAR-γ mRNA expression might depend on a decreased DBP protein.
Our previous study showed that HDACs inhibition increased the mRNA expressions of DBP and subsequent PPAR-γ1sv, the homolog of human novel PPAR-γ, in epididymal adipose tissue in ob/ob mice, but not in the control animals (C57BL/6JHamSlc).10 The following findings were also obtained by HDACs inhibition in ob/ob mice: (i) increased proportion of smaller size of preadipocytes in epididymal adipose tissue, which might be mediated through the action of elevated PPAR-γ1sv protein12; (ii) elevated plasma adiponectin concentration, which is mainly secreted by small size adipose tissue24; (iii) improvement of peripheral insulin signal transduction measured by phosphorylation of insulin-receptor substrate 1 (IRS-1) and AKT in muscle by the elevated plasma adiponectin concentration.25 In this study, patients with DM2 were treated with antidiabetic drugs including sulfonylureas and DPP-4 inhibitors until 1 day before surgery. Because these antidiabetic drugs are reported to elevate plasma adiponectin concentration,26–28 metabolic variables such as adiponectin, leptin, blood glucose and HbA1c were not measured in the present study. Therefore, an influence of reduced H3K9 acetylation at the promoter regions of Dbp in visceral adipose tissue on glucose metabolism could not be evaluated. Further studies including patients with DM2 without any antidiabetic drugs are needed to evaluate a potential influence of a reduced DBP protein on glucose metabolism.
Subcutaneous adipose tissue represents the normal physiological buffer for excess energy intake with limited energy expenditure. It acts as a metabolic sink where excess free fatty acids and glycerol are stored as triglycerides.29 Previous study reported that clock genes expressions are decreased in subcutaneous adipose tissue in patients with obesity having DM2,30 but other study showed that clock genes expressions are maintained in these patients.31 Omental and mesenteric adipose tissue are two major components of visceral fat. Their profiles are summarized as13: (i) omentum is the fatty apron attached to the greater curvature of the stomach, and mesentery is the fat containing the arteries and veins supplying the small and large intestine and (ii) visceral tissue adipocytes are more metabolically active, more sensitive to lipolysis and more insulin resistance than subcutaneous adipocytes. Previous study in patients without DM2 showed that the mRNA expressions of CLOCK, BMAL1, PER1, PER2 and CRY1 in omental adipose tissue were similar between obese and lean.32 The present study added data indicating that DM2 might not alter the expressions of these clock genes in omental and mesenteric adipose tissue.
To our knowledge, data concerning a potential different role of omental and mesenteric adipose tissue in insulin resistance of DM2 are largely lacking due to the difficulty in obtaining these tissue.33 34 In addition, these two studies provided diverse findings about the role of mesenteric33 and omentum34 adipose tissue in insulin resistance. In this study, the mRNA expressions of DBP and subsequent novel PPAR-γ in the patients with DM2 were significantly lower in the omental adipose tissue than in the mesenteric adipose tissue. On the other hand, Yang et al reported that PPAR-γ mRNA expression was upregulated in the mesenteric adipose tissue in patients with obesity having DM2.14 However, obesity per se is reported to elevate the PPAR-γ mRNA expression in visceral adipose tissue,32 which might partially explain the elevated expression of PPAR-γ mRNA in the omental adipose tissue in the study by Yang et al (mean BMI 44.9 kg/m2). Patients with obesity were not included in this study (mean BMI 24.0 kg/m2). Therefore, a big difference in BMI between our study patients and patients in the study by Yang et al might cause diverse influence on PPAR-γ mRNA expression in the omental adipose tissue.
Limitations
Gender differences are reported in chrono-physiological35 and chrono-pharmacological36 profiles in human subjects. In this study, the ratio of male/female in the DM2-gastric cancer group is larger (but not significantly) than that in the non-DM-gastric cancer group. Therefore, gender differences cannot be excluded as the cause of the significant difference between the DM2 and non-DM groups for the omental fat.
There were no significant seasonal differences in the number of patients enrolled in this study between the non-DM and DM2 groups. However, season would change the length of day light that a patient might be exposed and this would influence circadian gene expression profiles.
It is well-known that impaired adiponectin secretion is involved in the pathogenesis of insulin resistance, which leads to hyperinsulinemia. Serum insulin did not elevate in the patients with DM2 in this study. Therefore, a study involving patients with hyperinsulinemia is warranted for further evaluation of Dbp expression in visceral adipose tissue in patients with DM2.
Eating patterns may be strongly altered in patients with gastric cancer, which were not controlled before surgery in this study. Because a high-fat diet is reported to cause a phase advance in the mRNA expression of DBP,37 a difference in eating pattern in each patient with gastric cancer might affect Dbp gene expression in omental adipose tissue. Further study involving patients with gastric cancer under a control of eating pattern is needed.
Patients with cancer may lose body weight which may affect adipose tissue gene expression, and they may also undergo various diagnostic procedures. Therefore, whether the present findings also apply to patients with DM2 without cancer remains to be determined.
Clock genes mRNA expression showed a profound diurnal rhythm in subcutaneous adipose tissue in patients with DM2.30 To avoid the potential diurnal rhythm of clock genes in visceral adipose tissue, samples were obtained within relatively narrow o’clock time range (10:40–13:40) in this study. However, it remains to be determined whether the differences in DBP mRNA expression between DM2 group and the control group also persist throughout the circadian cycle.
In summary, the present study showed that H3K9 acetylation at the promoter regions of Dbp in omental adipose tissue was significantly reduced in patients with DM2. Consequently, mRNA expressions of DBP and novel PPAR-γ were significantly lower in patients with DM2. These changes were not detected in mesenteric adipose tissue. Because adiponectin secretion is reported to be enhanced through the PPAR-γ-related mechanism, this study supports the hypothesis that omental adipose tissue is involved in the mechanism of DM2.
References
Footnotes
Contributors Designed the research: KU and AF. Performed the laboratory experiments: KU, CS, HKi and KS. Performed the surgery: HH and YH. Dissected the tissues: HKa and AT. Analyzed data: KU, CS and HKi. Wrote the manuscript: KU, CY, YI and AF.
Funding This study was supported by Japan Society for the Promotion of Science; Grant-in-Aid for Young Scientists (B) (16K18954 to KU) and Fund for the Promotion of Joint International Research (Fostering Joint International Research) (17KK0198 to KU).
Competing interests None declared.
Patient consent for publication Not required.
Ethics approval The protocol of this study was reviewed and approved by the Ethics Review Board of Jichi Medical University (No. 18-hen29, Tochigi, Japan). Written informed consent was obtained from each patient before surgery.
Provenance and peer review Not commissioned; externally peer reviewed.
Data availability statement All data relevant to the study are included in the article or uploaded as supplementary information.