Article Text

Abstract
Introduction Lutein is a carotenoid whose protective effects in the retina have been reported in various studies. The effect of lutein has not been reported in the retina of the Ins2Akita/+ mouse, a well-characterized genetic model for diabetic retinopathy (DR) in which the etiology of diabetes is better defined than the chemically induced diabetes. The objective of the present study is to investigate the effect of long-term administration of lutein in early stages of DR using the Ins2Akita/+ mouse.
Research design and methods Heterozygous male Ins2Akita/+ and age-matched wild-type mice were used. Lutein was administered to the mice in drinking water starting 6 weeks old daily until analysis at 4.5, 6.5 or 9 months of age. Plain water served as non-treatment control. Microglia were immunostained with ionized calcium-binding adapter molecule 1 (Iba-1) and cluster of differentiation 68 (CD68) in retinal flat-mounts. Vascular endothelial growth factor (VEGF) level in the retina was assessed by enzyme-linked immunosorbent assay (ELISA). Vascular permeability was analyzed in retinal flat-mounts after fluorescein isothiocyanate (FITC)-dextran perfusion. Retinal occludin expression was assessed via Western blots. Retinal function was examined by electroretinography (ERG).
Results Increased microglial reactivity was detected in the Ins2Akita/+ mouse retina and was suppressed by lutein. Lutein administration also reduced the upregulation of VEGF in the Ins2Akita/+ mouse retina. Increased vascular leakage and decreased occludin expression were observed in the Ins2Akita/+ mouse retina, and these alterations were attenuated by lutein treatment. ERG recordings showed reduced a-wave and b-wave amplitudes in the Ins2Akita/+ mice. With lutein treatment, the ERG deficits were significantly alleviated.
Conclusions We showed beneficial effects of long-term lutein administration in the Ins2Akita/+ mouse retina, including suppression of retinal inflammation, protection of retinal vasculature and preservation of retinal function. These results point to lutein’s potential as a long-term therapeutic intervention for prevention of inflammation and retinal degeneration in patients with early DR.
- anti-inflammatory agents
- antioxidants
- blood-retinal Barrier
- carotenoids
This is an open access article distributed in accordance with the Creative Commons Attribution Non Commercial (CC BY-NC 4.0) license, which permits others to distribute, remix, adapt, build upon this work non-commercially, and license their derivative works on different terms, provided the original work is properly cited, appropriate credit is given, any changes made indicated, and the use is non-commercial. See: http://creativecommons.org/licenses/by-nc/4.0/.
Statistics from Altmetric.com
Significance of this study
What is already known about this subject?
Microglial reactivity is increased in the retinas of the Ins2Akita/+ mice.
Impairment of retinal function was observed in the Ins2Akita/+ mice.
What are the new findings?
Lutein treatment suppressed inflammatory responses (microglial reactivity and vascular endothelial growth factor upregulation) and attenuated retinal vascular leakage in the retinas of the Ins2Akita/+ mice.
Lutein treatment protected retinas from functional impairment in the Ins2Akita/+ mice.
Long-term application of lutein at 4.2 mg/kg/day (equivalent human dose 20 mg/day) provided comparable beneficial effects to the higher dose 8.4 mg/kg/day (equivalent human dose 40 mg/day) in the Ins2Akita/+ mice.
How might these results change the focus of research or clinical practice?
Our study may provide a rationale for long-term application of lutein in large clinical trials in subjects with early stages of diabetic retinopathy.
Introduction
Diabetic retinopathy (DR) is the most common microvascular complication of diabetes mellitus. As a sight-threatening condition, DR remains the main cause of new blindness in working-age populations. One-third of 422 million individuals with diabetes around the world are estimated to suffer from DR, and the prevalence is expected to continue to rise.1 DR falls into two stages: non-proliferative diabetic retinopathy (NPDR) and proliferative diabetic retinopathy (PDR). NPDR represents the early stage of DR, in which the patients are typically asymptomatic. The major cause of vision loss in patients with NPDR is diabetic macular edema (DME) resulting from intraretinal accumulation of fluid from leaky blood vessels.2 As the disease evolves into PDR, which is the late stage of DR characterized by neovascularization, patients will be at high risk of serious vision loss induced by vitreous hemorrhage or tractional retinal detachment.
Current treatments for DR include anti-vascular endothelial growth factor (VEGF) injection, laser photocoagulation and intravitreal steroid therapy. These treatments target directly on the sight-threatening retinal vascular injuries and are typically employed at relatively advanced stages. However, the overall response rate is less than 50%.3 Recently, evidence suggest that DR is not only a disorder of retinal vasculopathy.4 5 Inflammation and retinal neuronal defects were detected in the initial stages of DR, prior to the occurrence of retinal vascular changes.4 5 This indicated that inflammation and retinal neurodegeneration might be implicated in DR independent of the diabetes-induced vasculopathy. This emphasizes an urgent need for the development of novel interventions to delay or halt DR progression in the early stage. Since long-term use is expected, an acceptable safety profile is of particular concern in the development of any new early intervention.
Lutein is a naturally occurring carotenoid pigment uniquely found in the human retina. It is a well-known antioxidant which possesses potent reactive oxygen species (ROS) scavenging capacity. As a generally recognized as safe (GRAS) molecule, lutein possesses a high safety profile.6 After being absorbed from food sources such as eggs, spinach and kale, lutein is specifically accumulated into human lens and retina from the blood.7 8 Steroidogenic acute regulatory domain protein 3 (StARD3) has been recognized as a lutein-binding protein.9 Lutein is proposed to help maintain eye health through potent antioxidant and blue light-absorbing properties.10 Both in vitro and in vivo studies demonstrated that lutein might protect the retina in pathological conditions via antiapoptotic and antioxidant activities.11 12 Moreover, lutein was reported to have an anti-inflammatory effect in the retina following retinal ischemia/reperfusion injury.13 Large-scale clinical studies have been carried out to investigate lutein’s effect in age-related macular degeneration (AMD), an aging eye disease related to oxidative stress and inflammation.14 15 Results from the Age-related Eye Disease Study 2 (AREDS2) suggested an inverse association between lutein consumption and risk of developing advanced AMD.14 Consistent evidence was provided in another clinical study, the Lutein Antioxidant Supplementation Trial, which further showed lutein’s effectiveness in improving visual function in patients with AMD.15 Similar to AMD, DR is an eye disorder associated with oxidative stress, inflammation as well as retinal neurodegeneration. Therefore, lutein may be a promising candidate as an agent or adjunct therapy in early DR intervention. The effect of lutein in the Ins2Akita/+ mouse, a well-characterized genetic model for DR, has not been reported before. The goal of the present study was to investigate whether long-term lutein supplementation has beneficial effects in early DR using the Ins2Akita/+ mouse model.
The etiology of diabetes in the Ins2Akita/+ mouse is better-defined than the chemically induced diabetes.16 The Ins2Akita/+ mice carry a spontaneous mutation (single base-pair substitution) in the insulin 2 (Ins2) gene, resulting in proinsulin protein misfolding and subsequent beta cell failure, eventually leading to spontaneous insulin-dependent diabetes.17 The male heterozygous Ins2Akita/+ mice develop hyperglycemia from around 4 weeks of age and remain stable thereafter. They exhibit typical features of early DR including vascular abnormalities, retinal neurodegeneration as well as glial dysfunction.18
Methods and materials
Animals and diets
C57BL/6J Ins2Akita/+ heterozygote mice (Stock No: 003548) were obtained from the Jackson Laboratory (Bar Harbor, Maine, USA). Animals were bred and housed in our specific pathogen free facility with a 12/12 hour light/dark cycle. All animals had free access to water and a standard chow diet containing no lutein (PicoLab Rodent 20 #5053, Purina Mills, St Louis, Missouri, USA). Animals were monitored daily for general health. Genotype of Ins2Akita/+ mice was confirmed using the protocol provided by the Jackson Laboratory (https://www.jax.org/Protocol?stockNumber=003548&protocolID=176). Hyperglycemia in Ins2Akita/+ mice was confirmed by blood glucose level higher than 250 mg/dL, using a Bayer Contour blood glucose meter (Bayer HealthCare, Diabetes Care, Tarrytown, New York, USA). Only heterozygous male Ins2Akita/+ mice were included in this study. All animal experiments adhered to the Association of Research in Vision and Ophthalmology Statement for the Use of Animals in Ophthalmic and Vision Research and were approved by the Committee on the Use of Live Animals in Teaching and Research of The University of Hong Kong (CULATR 4201–16).
Lutein treatment
A starch-based, water dispersible form of lutein (FloraGLO Lutein 10% CWS/S-TG, Kemin Industries, Des Moines, Iowa, USA) was administered to the mice in drinking water daily starting from 6 weeks of age until analysis. Three doses, 2.1, 4.2 and 8.4 mg/kg/day, were tested in a pilot study. The initial starting dose 2.1 mg/kg/day was converted from an oral human equivalent dose (HED), 10 mg/day, used in the AREDS214 using an interspecies allometric scaling method.19 Only the two higher doses, 4.2 and 8.4 mg/kg/day, were found to be protective on visual function by 6.5 months of age, which therefore were used in our subsequent experiments. The wild-type (WT) control group and untreated Ins2Akita/+ group received drinking water only. Amber-colored water bottles were used to avoid light exposure. Lutein was changed every 2 days and the concentration was adjusted based on water consumption and body weight. n≥5 mice per group were used in each experiment.
Immunostaining of retinal flat-mounts
Animals were sacrificed after euthanasia with pentobarbital overdose. Eyes were enucleated immediately and fixed in 4% (wt/vol.) paraformaldehyde (PFA) for 1 hour at room temperature. Following fixation, whole retinas were dissected and washed with 0.01 mol/L phosphate buffered saline (PBS) for 2–3 hours on a rotator. PBS were changed every 30 min. Retinas were then blocked for 1 hour in blocking buffer (10% (wt/vol.) bovine serum albumin (BSA) in 0.1% (vol./vol.) TBS-Tween-20). For primary antibody incubation, retinal samples were incubated in primary diluent overnight at 4°C with the following antibodies: rabbit anti-ionized calcium-binding adapter molecule 1 (Iba-1, 1:500, Wako Chemicals, Richmond, Virginia, USA) and rat anti-cluster of differentiation 68 (CD68, 1:500, Abcam, Cambridge, Massachusetts, USA). After six washes with PBS, retinas were incubated with AlexaFluor 488-conjugated or 555-conjugated secondary antibodies overnight at 4°C. Following washing, whole retinas were flat-mounted onto microscope slides with the ganglion cell side facing up. After air-drying, specimens were cover-slipped with mounting medium and were stored at −20°C. Images were taken with the Zeiss LSM 780 Inverted Confocal Microscope (Zeiss, Jena, Germany) with a 20×objective and were analyzed blindly.
Retro-orbital injection
Retro-orbital injection of dextran (FD70S, Sigma-Aldrich, St Louis, Missouri, USA) conjugated with fluorescein isothiocyanate (FITC-dextran) was performed as previously described.20 Animals were anesthetized with a combination of ketamine/xylazine (100 and 10 mg/kg body weight, respectively) via intraperitoneal injection. Ten milligrams of FITC-dextran per mouse dissolved in 0.2 mL 0.01 mol/L PBS was injected into the retro-orbital venous sinus. Ten minutes later, animals were sacrificed by pentobarbital overdose and their eyes were enucleated immediately. Eyeballs were fixed for 1 hour at room temperature in 4% (wt/vol.) PFA while kept in darkness. Subsequently, the whole retina was dissected and flat-mounted onto a microscopic slide. The slides were stored at −20°C in the dark until imaging using the Zeiss LSM 780 Inverted Confocal Microscope (Zeiss, Jena, Germany) with a 5× objective. Tile scanning was employed to image the entire retina. Zeiss Zen software was used for image processing and the images were analyzed blindly.
ELISA
VEGF level in the mouse retina was assessed using a Mouse VEGF Quantikine enzyme-linked immunosorbent assay (ELISA) Kit (MMV00, R&D systems, Minneapolis, Minnesota, USA). At 6.5 and 9 months of age, mouse retinas were collected and processed following the manufacturer’s instructions. Briefly, retinas were homogenized ultrasonically in 0.01 mol/L PBS containing protease inhibitors in an ice-water bath. Samples were then stored at −80°C overnight and thawed out on ice. Following two freeze-thaw cycles, the samples were centrifuged at 5000 g for 5 min at 4°C. The bicinchoninic acid (BCA) protein assay was performed for determination of total protein concentration. Absorbance of each well was measured at 450 nm using a multimode microplate reader (BioTek Instruments, Winooski, Vermont, USA). Inclusion of a standard curve (7.8–500 pg/mL) allowed determination of VEGF level in each sample.
Western blot analysis
For protein isolation, retinal samples underwent ultrasonic homogenization for 20 min in ice-cold radioimmunoprecipitation assay (RIPA) lysis buffer containing protease inhibitors. After determination of protein concentration by the BCA assay, 20 ug protein from each sample was separated by 10% (wt/vol.) sodium dodecyl sulphate-polyacrylamide gel electrophoresis (SDS-PAGE) gels and then transferred to a polyvinylidene fluoride (PVDF) membrane. Following blocking with 5% (wt/vol.) fat-free milk, membranes were incubated overnight at 4°C with the following primary antibodies: rabbit anti-occludin antibody (1:1000, 71–1500, Thermo Fisher Scientific, Waltham, Massachusetts, USA) and mouse anti-β-actin antibody (1:5000, Chemicon, Temecula, California, USA). After washing in Tris-buffered saline with Tween 20 (TBST), membranes were incubated with horseradish peroxidase-conjugated secondary antibodies (Vector Laboratories, Inc., Burlingame, California, USA). Protein bands were visualized with WesternBright chemiluminescent detection reagent (Advansta Inc., San Jose, California, USA). The band intensities were analyzed using Image J and were normalized with β-actin.
Electroretinogram
Electroretinogram (ERG) was performed as previously described.13 Animals were dark-adapted overnight and then anesthetized under dim red light with the ketamine/xylazine as described above. One per cent (wt/vol.) Tropicamide (Mydriacyl, Alcon-Couvreur, Puurs, Belgium) was applied topically to dilate the pupils. Moisturizing eye drops (Tears Naturale II, Alcon-Couvreur, Puurs, Belgium) were used to keep the cornea lubricated. Topical anesthetic proparacaine hydrochloride (0.5% (wt/vol.), Alcaine, Alcon-Couvreur, Puurs, Belgium) was applied to the cornea as needed. For ERG signal recording, gold ring electrodes were placed on the cornea. Subcutaneous needle electrodes were inserted at the dorsal midline between two ears and the tail serving as the negative and ground references, respectively. Scotopic ERG responses were recorded using the Color Dome Ganzfeld System (Diagnosys Espion system, Diagnosys, Lowell, Massachusetts, USA) at light intensities of 3.0 and 10.0 cd s/m2, respectively. Amplitudes of a-wave and b-wave were analyzed.
Statistical analysis
Statistical comparisons among multiple groups were performed by one-way analysis of variance (ANOVA) followed by posthoc Tukey’s multiple comparison test (Prism 5; GraphPad Software, San Diego, California, USA). Mean ± standard error of the mean (SEM) was presented. P<0.05 was considered statistically significant.
Results
Suppression of microglial reactivity by lutein treatment in the retinas of Ins2Akita/+ mice
Microglia are a major neuroglial cell type in the retina. On activation, microglial cells undergo conversion from “surveillant” phenotype (ramified) into a “reactive” (ameboid) state. In this study, we evaluated the immunoreactivity of microglia on retinal flat-mounts using two markers: Iba-1 which stains both ramified and ameboid microglia, and CD68 which labels ameboid microglia. As shown in figure 1, with anti-Iba-1 antibody (green), ramified microglia were identified with small round cell body with highly ramified processes (white arrow heads). In contrast, ameboid microglia exhibited a larger cell body with shortened processes which were double stained with anti-Iba-1 and anti-CD68 (red) antibodies (white arrows). After counting the number of reactive microglia (doubly stained), we found that at the age of 4.5 months, the number of Iba-1 and CD68 double immunopositive microglia was significantly higher in the Ins2Akita/+ mouse retina when compared with that in the WT control group, suggesting increased microglial immunoreactivity in the diabetic mouse retina. However, with lutein treatment, microglial immunoreactivity was markedly suppressed when compared with the untreated group (figure 1). No significant difference in body weight or glucose level was observed between the lutein-treated and untreated groups (data not shown).

Suppression of microglial reactivity by lutein treatment in the retinas of the Ins2Akita/+ mice. (A) Representative images of the retinal flat-mounts immunostained with CD68 and Iba-1 from the wild-type (a–c), untreated Ins2Akita/+ (d–f), lutein-treated Ins2Akita/+ (4.2 mg/kg/day, Akita+L4.2) (g–i) and lutein-treated Ins2Akita/+ (8.4 mg/kg/day, Akita+L8.4) (j–l) mice at 4.5 months of age. Photomicrographs showed CD68-positive reactive microglia (red, white arrows) and Iba-1-positive total microglia (green, white arrow heads), respectively, together with merged images. Scale bar=100 µm. (B) Quantitative results of number of reactive microglia in the four experimental groups. n=6. Data are presented as mean±SEM; One-way ANOVA followed by Tukey’s multiple comparison test. **P<0.01, ****p<0.0001. ANOVA, analysis of variance; CD68, cluster of differentiation 68; Iba-1, ionized calcium-binding adapter molecule 1.
Lutein treatment alleviated the upregulation of VEGF in the retinas of the Ins2Akita/+ mice
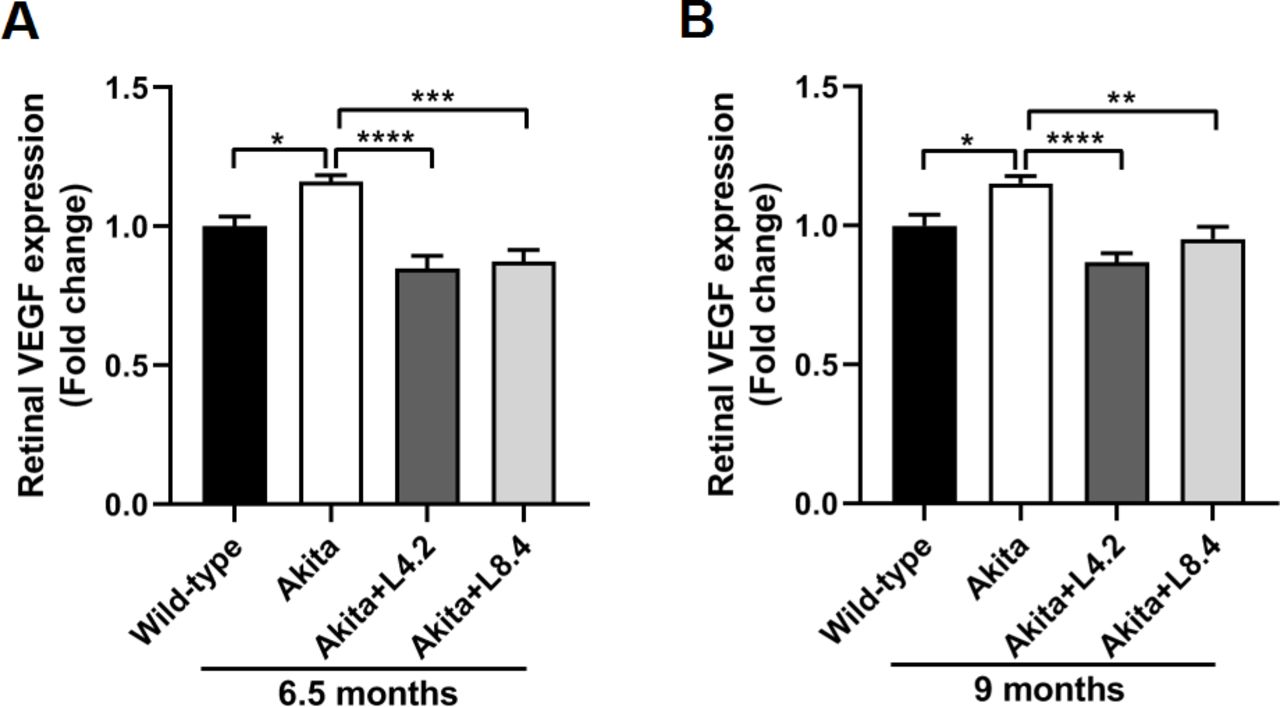
VEGF-A (hereafter refer to as VEGF) has been implicated in both NPDR and PDR. Its critical role in regulating retinal vascular permeability and angiogenesis has been extensively described. To examine whether lutein can modulate VEGF level in the Ins2Akita/+ mice, we evaluated the expression of VEGF in retinas by ELISA. Our data showed that VEGF protein level in the retinas of Ins2Akita/+ mice was significantly higher than that in the WT mice at both 6.5 (p<0.05) and 9 (p<0.05) months of age, suggesting an upregulation of VEGF expression in the diabetic mouse retina. With lutein treatment, the increased VEGF level in the Ins2Akita/+ mouse retina was downregulated at both doses (4.2 and 8.4 mg/kg/day) when compared with that in the untreated mouse retina (figure 2).
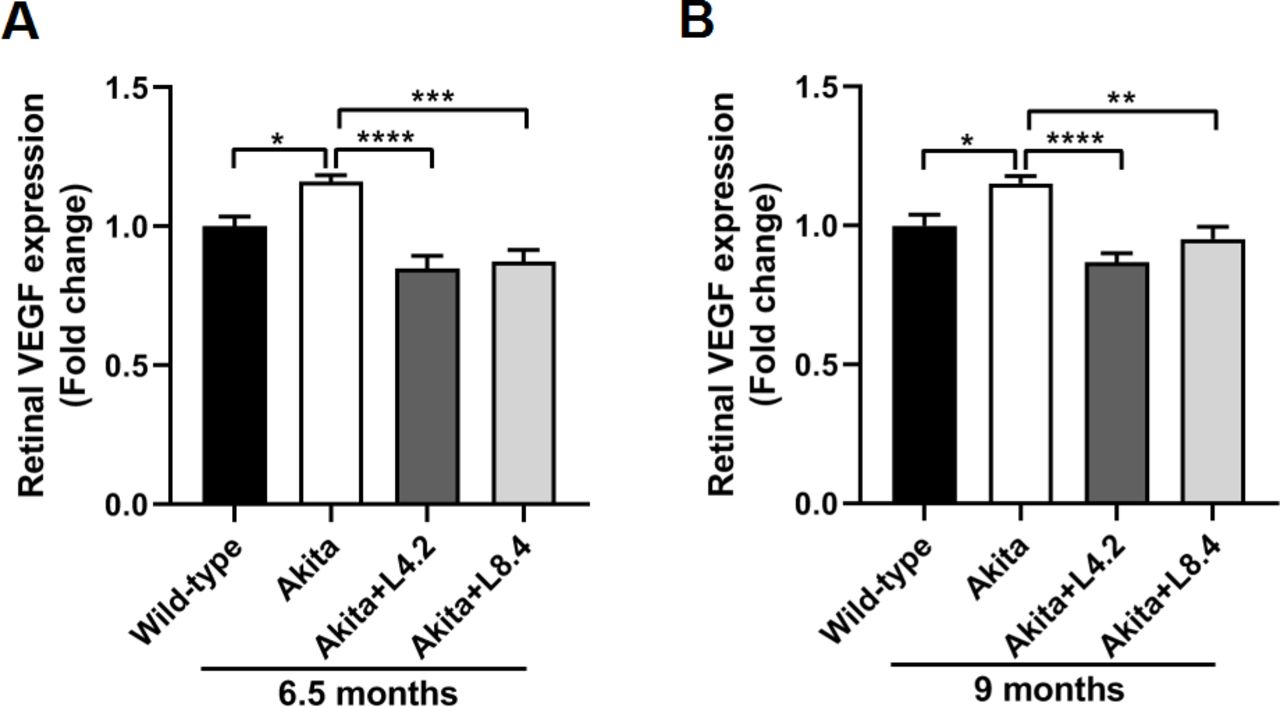
Lutein treatment alleviated the upregulation of VEGF in the retinas of the Ins2Akita/+ mice. (A) ELISA results showing retinal VEGF protein expression levels in the wild-type, untreated Ins2Akita/+, lutein-treated Ins2Akita/+ (4.2 mg/kg/day, Akita+L4.2) and lutein-treated Ins2Akita/+ (8.4 mg/kg/day, Akita+L8.4) mice at 6.5 months of age. (B) ELISA results showing retinal VEGF levels in the four groups at 9 months of age. n=5–8. Data are presented as mean±SEM; One-way ANOVA followed by Tukey’s multiple comparison test. *P<0.05, **p<0.01, ***p<0.001, ****p<0.0001. ANOVA, analysis of variance; ELISA, enzyme-linked immunosorbent assay; VEGF, vascular endothelial growth factor.
Reduction of retinal vascular leakage by lutein in the retinas of the Ins2Akita/+ mice
FITC-dextran extravasation method was employed to assess retinal vascular permeability, an indicator of blood-retinal barrier (BRB) integrity. After perfusion with retro-orbitally injected FITC-dextran, leakage of FITC-dextran from the retinal vasculature was evaluated in the flat-mounted retina. As shown in figure 3, retinal vascular leakage in the retinas of Ins2Akita/+ mice appeared to be increased when compared with that in the WT control. The number of leakage sites in the entire retina was counted and compared among the four experimental groups. This revealed a significant increase in the number of leakage sites in the 9-month-old Ins2Akita/+ mouse retina when compared with the age-matched WT controls (p<0.01), indicating a disruption of BRB in the 9 month old Ins2Akita/+ mice. However, with lutein treatment (4.2 and 8.4 mg/kg/day), the number of FITC-dextran extravasation and hence vascular leakage in the Ins2Akita/+ mouse retina was decreased when compared with the untreated mice (figure 3, p<0.05 and p<0.01).

Reduction of retinal vascular leakage by lutein treatment in the retinas of Ins2Akita/+ mice. (A) Images of FITC-dextran-perfused retinal flat-mounts from the wild-type, untreated Ins2Akita/+, lutein-treated Ins2Akita/+ (4.2 mg/kg/day, Akita+L4.2) and lutein-treated Ins2Akita/+ (8.4 mg/kg/day, Akita+L8.4) mice at 9 months of age. Representative images of the entire retinal flat-mounts from the four groups are shown (a–d). Images taken from the corresponding regions in a–d (white rectangles) at higher magnification are shown in e–f. Extravasation of FITC-dextran resulting from retinal vascular damage is shown (white arrows). Scale bars: a–d=500 µm, e–h=100 µm. (B) Quantitative results of number of leakage sites in retinas from four groups. n=7–8. Data are presented as mean±SEM; One-way ANOVA followed by Tukey’s multiple comparison test. *P<0.05, **p<0.01. ANOVA, analysis of variance; FITC, fluorescein isothiocyanate.
Alleviation of tight junction protein disruption by lutein in the retinas of Ins2Akita/+ mice
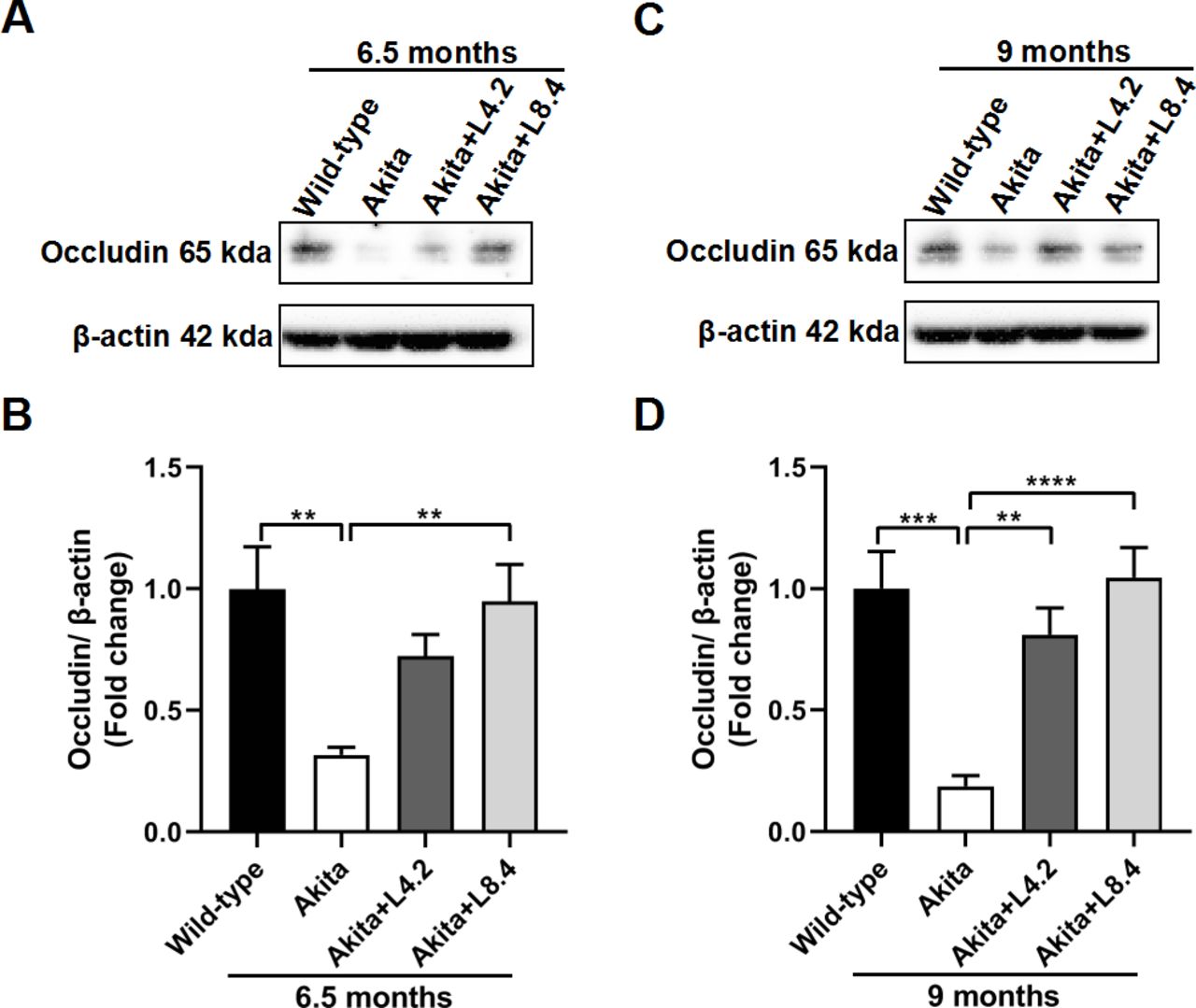
To further investigate the role of lutein on retinal vascular integrity, we analyzed the expression of a tight junction protein, occludin, by Western blots. In retinas of the untreated Ins2Akita/+ mice, occludin protein content was markedly decreased when compared with that in the WT mice at both 6.5 (p<0.01) and 9 (p<0.001) months of age. This may partially explain the breakdown of BRB and increase of retinal vascular permeability in the untreated Ins2Akita/+ mice as observed above. After treatment with lutein, the reduction in occludin expression in the Ins2Akita/+ mice was alleviated when compared with the untreated group at both 6.5 and 9 months of age (figure 4).

Alleviation of tight junction protein disruption by lutein treatment in the retinas of the Ins2Akita/+ mice. (A,B) Western blot results showing occludin expression levels in the wild-type, untreated Ins2Akita/+, lutein-treated Ins2Akita/+ (4.2 mg/kg/day, Akita+L4.2) and lutein-treated Ins2Akita/+ (8.4 mg/kg/day, Akita+L8.4) mice at 6.5 months of age. (C,D) Western blot results showing occludin expression levels in the four experimental groups at 9 months of age. Occludin expression level in each group was normalized with β-actin. Fold changes relative to wild-type control are shown. n=6–7. Data are presented as mean±SEM. One-way ANOVA followed by Tukey’s multiple comparison test. **P<0.01, ***p<0.001. ****p<0.0001. ANOVA, analysis of variance.
Preservation of retinal function by lutein in the Ins2Akita/+ mice
ERG test is a sensitive method in assessing retinal neuronal integrity. In Ins2Akita/+ mice, a decrease of both scotopic a-wave and b-wave amplitudes was detected when compared with the WT control at 6.5 and 9 months of age (figure 5), suggesting a deterioration of retinal neuronal function caused by diabetes. However, with lutein treatment, the decrease of ERG responses in the Ins2Akita/+ mice was significantly ameliorated at both 6.5 and 9 months of age (figure 5).

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Preservation of retinal function by lutein treatment in the Ins2Akita/+ mice. (A–C) ERG responses in the wild-type, untreated Ins2Akita/+, lutein-treated Ins2Akita/+ (4.2 mg/kg/day, Akita+L4.2) and lutein-treated Ins2Akita/+ (8.4 mg/kg/day, Akita+L8.4) mice at 6.5 months of age. Representative ERG waveforms (flash intensity 10 cd s/m2) from the four groups are shown (A). Comparison of a-wave and b-wave amplitudes in the four groups at light intensities of 3.0 (B) and 10.0 (C) cd s/m2, respectively, is shown. n=17–25. (D–F) ERG responses in the four experimental groups at 9 months of age. Representative ERG waveforms (flash intensity 10 cd s/m2) from the four groups are shown (D). Comparison of a-wave and b-wave amplitudes in the four groups at light intensities of 3.0 (E) and 10.0 (F) cd s/m2, respectively, is shown. n=9–11. One-way ANOVA followed by Tukey’s multiple comparison test. Data are presented as mean±SEM. *P<0.05, **p<0.01, ***p<0.001. ****p<0.0001. ANOVA, analysis of variance; ERG, electroretinogram; WT, wild-type.
Discussion
Currently, anti-VEGF injection remains the mainstay of DR therapy. However, requirements for repeated and long-term use can be accompanied by devastating ocular side effects given VEGF’s role in retinal development and function.21 Lutein is an essential nutrient whose long-term safety has been well demonstrated in both animal and human studies with no reported side effects.7 22 This is the first study reporting the protective effects of long-term lutein administration in the Ins2Akita/+ mice, a type 1 diabetes model which has been shown to mimic early DR with multiple advantages. Our data on the regulatory effect of lutein on retinal microglial reactivity, VEGF expression and retinal neuronal function in Ins2Akita/+ mice indicate that lutein may serve as an early intervention for DR, in favor of targeting inflammation and/or neurodegeneration preceding the microvascular changes in the diabetic retina in the early stages.
Although the exact mechanisms underlying the pathophysiology of DR remain inconclusive, inflammation has gained increasing interest as a contributory pathway in DR.23–25 Microglial activation has been proposed to play a central role in diabetes-induced retinal inflammation, by releasing proinflammatory cytokines including tumor necrosis factor α (TNF-α), Interleukin-1β (IL-1β), IL-6 and so on.26–28 ROS, as a secondary messenger, has been shown to be a key factor mediating proinflammatory responses in activated microglia and continued microglial activation.29 30 In practice, corticosteroids, as potent anti-inflammatory agents, have demonstrated remarkable efficacy in treating DME.31 However, long-term utilization of intravitreal steroids is of great concern given the high-incidence of sight-threatening ocular side effects.31 In our study, we demonstrated a suppressive effect of lutein on the activation of microglia in the Ins2Akita/+ mice. This result is consistent with findings in an in vitro study, wherein lutein showed inhibitory effects on microglial activation, release of TNF-α, IL-1β as well as ROS production.28 Hence, we speculate that lutein may protect the diabetic retina from inflammatory damage through its ROS scavenging capacity. Although microglial reactivity was only examined at 4.5 months of age in our study, previous study has shown involvement of microglial activation throughout the disease course of DR.32 Therefore, early and long-term intervention of microglial reactivity with lutein treatment may provide favorable clinical outcome in DR treatment.
VEGF is a growth factor essential in both retinal vascular permeability and neovascularization. VEGF expression is markedly elevated in the vitreous fluid and aqueous humor of patients with DR, and its level is correlated with disease severity.33 Several pathways have been implicated in the upregulation of VEGF during DR, including hypoxia, production of advanced glycation end-products, oxidative stress and inflammatory activities.34 35 Blockade of the VEGF pathway using high affinity neutralizing antibodies has exhibited profound efficacy in both DME and PDR. However, depletion of VEGF may produce unexpected side effect given its role in normal retinal development and neuronal function.21 In vivo evidence revealed that sustained administration of anti-VEGF agents may lead to impairment of retinal cells and function.21 36 Therefore, neutralizing VEGF to a desired level without disrupting its physiological function in the retina is of great importance in practice. In our study, lutein treatment potently prevented the increase of VEGF in the retinas of the Ins2Akita/+ mice. Unlike the anti-VEGF agents who directly neutralize retinal VEGF, lutein may help to reduce pathologic VEGF levels by suppressing augmented oxidative stress and inflammatory activities.34 37 Moreover, evidence from in vitro studies showed that lutein could partially prevent hypoxia-induced apoptosis and downregulate the expression of hypoxia inducible factor-1α (HIF-1α) following hypoxic insult, revealing lutein’s potential in suppressing hypoxia-induced injuries. As HIF-1α regulates VEGF level, lutein may inhibit pathologic VEGF expression by targeting the hypoxia signaling pathways in the diabetic retina.
We also investigated the effect of long-term lutein treatment on integrity of the BRB. BRB plays a crucial role in maintaining retinal homeostasis. Breakdown of BRB is a hallmark feature of DR which leads to abnormal leakage of fluid and blood components from the retinal vasculature into the retina. VEGF has been recognized as a major contributor to BRB impairment in the diabetic retina.33 In our study, increased BRB permeability was detected in the 9-month-old Ins2Akita/+ mice, as indicated by increased extravasation of FITC-dextran dye in the diabetic retina when compared with that in the age-matched WT control. This indicated BRB breakdown and leakage induced by hyperglycemia in the diabetic Ins2Akita/+ mice, which aligned with the increase in VEGF level. With long-term lutein treatment, FITC-dextran extravasation in the Ins2Akita/+ mice was significantly decreased when compared with that in the untreated diabetic mice. The restoration of BRB function by lutein may be, at least partially, ascribed to neutralization of VEGF as described above.
Occludin is a key factor in regulating BRB integrity. It is an integral tight junction transmembrane protein which constitutes a core element of the BRB.38 Studies showed that occludin plays an essential role in regulating tight junction stability and BRB permeability. Dysregulation of occludin is a fundamental mechanism by which many pathological factors induce tight junction disruption.39 In DR, VEGF-induced BRB impairment has been attributed to alterations of occludin content and phosphorylation status.40 In addition, other proinflammatory factors such as TNF-α and IL-1β have also been shown to mediate BRB breakdown by downregulating occludin expression.41 In our study, a decrease in occludin content was detected in the retinas of 6.5-month-old and 9-month-old Ins2Akita/+ mice, further confirming a disruption of the BRB tight junctions. In Ins2Akita/+ mice receiving long-term lutein treatment, the expression of occludin in the retina was significantly increased when compared with that in the untreated diabetic mice. It is worth noting here that, while the VEGF levels were comparable in the two treatment groups (figure 2), occludin expression in the 8.4 mg/kg lutein-treated group appeared to be higher than that in the 4.2 mg/kg lutein-treated group. This indicates that in addition to VEGF neutralization, lutein may further increase occludin content through modulation of other occludin regulatory pathways, such as the matrix metalloproteinase-mediated pathway.37 42
Retinal neurodegeneration has been recognized as an independent pathogenic pathway during DR.5 Using ERG, functional impairment has been reported in different cell types in the diabetic retina, including photoreceptors (rods and cones), bipolar cells and amacrine cells.43 44 In our study, impairment of rod photoreceptors and second-order retinal neurons (ON-bipolar cells) were detected in the Ins2Akita/+ mice, as indicated by significant decreases in scotopic a-wave and b-wave amplitudes. These results are consistent with findings in a previous study, wherein reduced amplitudes of scotopic a-wave and b-wave were observed in the Ins2Akita/+ mice.43 The observed retinal functional deficits were significantly ameliorated by long-term lutein administration, suggesting a neuroprotective potential of lutein in the diabetic retina. Indeed, lutein is neuroprotective and increases retinal neuron survival in various injuries, including acute ischemia/reperfusion injury and retinal detachment.12 13 45 46 Lutein may protect retinal neurons from diabetes-induced damage through two mechanisms: (1) direct suppression of ROS-mediated neurotoxicity47 and (2) prevention of ischemic retinal damage by alleviating retinal vascular abnormalities.
Consistent with our findings, protective effects of lutein have been shown in chemically induced diabetes. However, in these animal studies, effect of lutein was tested in a relative short period of time or at doses higher than the reported safety dose in human (maximum: 40 mg/day).47 48 In our study, two doses of lutein (HED 20 and 40 mg/day, respectively) were used in the Ins2Akita/+ mice and both exhibited beneficial effects. Notably, no significant difference was observed between the efficacy of the high and low doses. Previous study showed that the absorption efficiency of lutein may decrease as lutein concentration increases;49 this may partially explain our observation. Forty mg/day is the highest dose of lutein used in human and was demonstrated to be safe in a 2-month study.50 Our data suggest that long-term application of lutein at 20 mg/day may provide comparable beneficial effects to the higher dose 40 mg/day.
Conclusion
Ins2Akita/+ mice exhibited stable hyperglycemia and are demonstrated to be a good model for the early stage of DR. The protective effect of long-term lutein feeding in the retinas of Ins2Akita/+ mice points to lutein’s role as a potential effective treatment for patients with early DR. Most importantly, lutein’s effect on retinal inflammation inhibition and retinal functional protection indicates that lutein may be an effective intervention targeting early pathways preceding microvascular injuries in DR. With substantial efficacy and long-term safety, lutein may serve as a reliable and promising agent for the early intervention of DR and delays its progression to the more advanced PDR when invasive treatments such as anti-VEGF injection, laser photocoagulation and intravitreal steroid therapy are required.
Acknowledgments
The authors thank Zhongjie Fu, PhD (Department of Ophthalmology, Boston Children’s Hospital/Harvard Medical School, Boston, Massachusetts, USA) for her helpful comments and suggestions in the preparation of the manuscript. We thank C. Peng (Kemin Industries(Zhuhai) Co., Ltd.) for providing FloraGLO lutein. The authors acknowledge the assistance of Faculty Core Facility, Li Ka Shing Faculty of Medicine, The University of Hong Kong.
References
Footnotes
Contributors ACYL conceived and designed the study. WW, KCT, TCN and KLSC performed the animal experiments. WW, TCN and RKG performed the molecular studies and analyzed and interpreted the data. WW and ACYL wrote and revised the manuscript. ACYL had primary responsibility for final content. All authors participated in discussions and approved the final manuscript.
Funding This research work is supported by the Health and Medical Research Fund, the Food and Health Bureau, The Government of the Hong Kong Special Administrative Region (04150746) and The University of Hong Kong Seed Funding Programme for Basic Research (201310159038).
Competing interests None declared.
Patient consent for publication Not required.
Provenance and peer review Not commissioned; externally peer reviewed.
Data availability statement All data relevant to the study are included in the article.