Differences in Pregnancy Metabolic Profiles and Their Determinants between White European and South Asian Women: Findings from the Born in Bradford Cohort

 , , ,
, , ,
Abstract
:1. Introduction
2. Results
2.1. Participant Characteristics
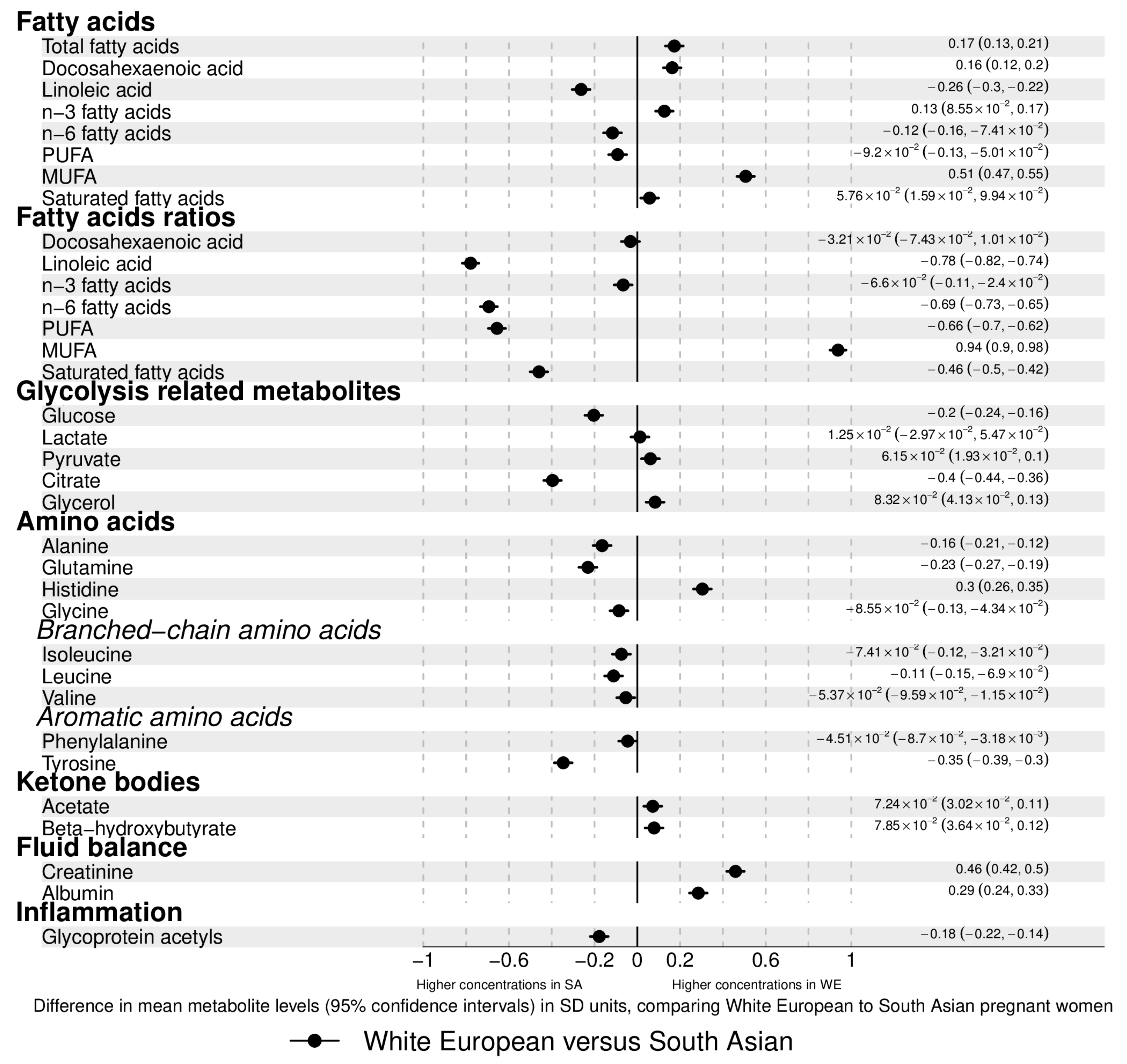
2.2. Ethnic Differences in Pregnancy Metabolic Profiles
2.3. Associations of Age, Education, and Parity with Gestational Metabolic Profiles in White European and South Asian Women
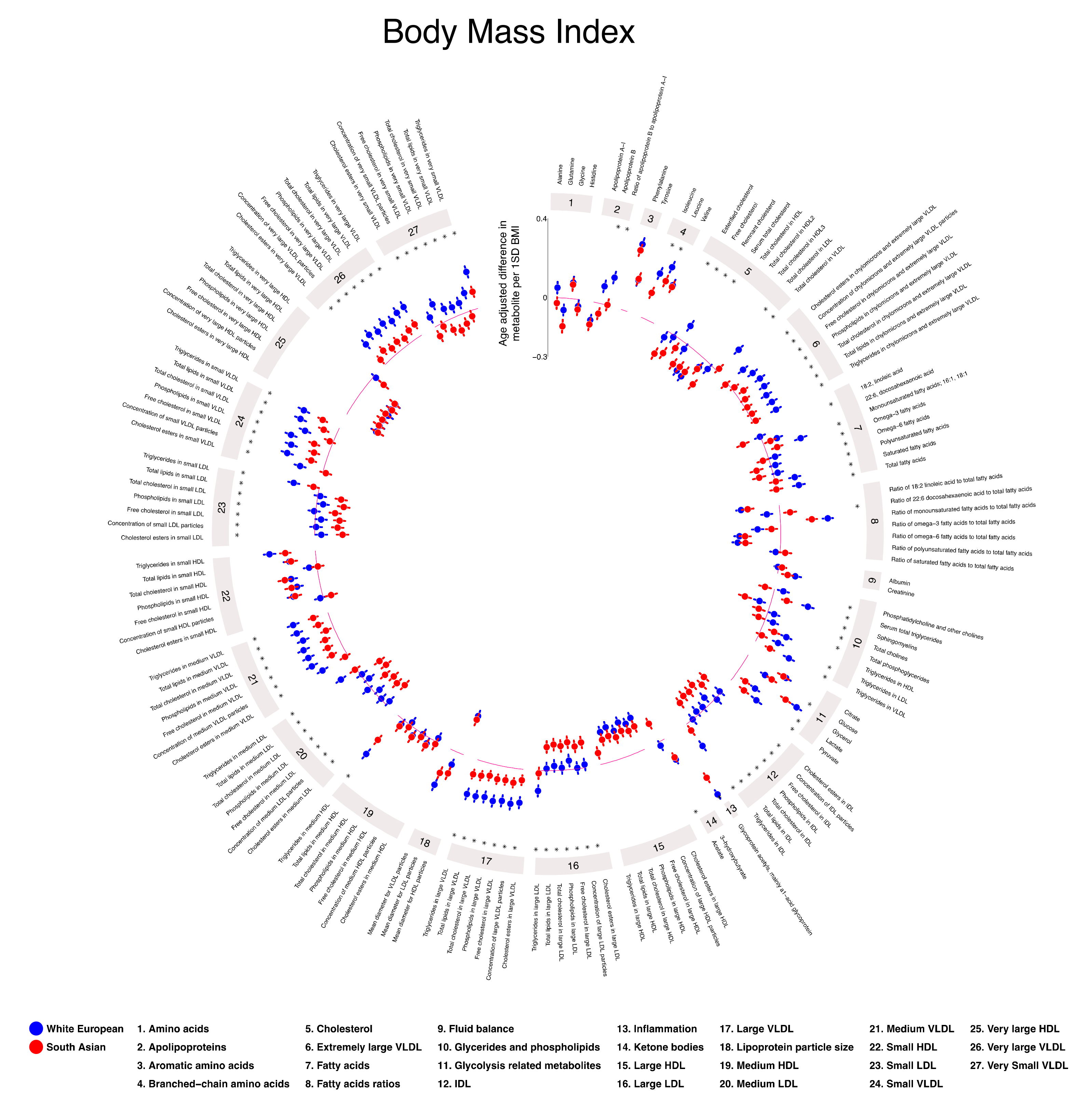
2.4. Associations of Height, BMI, and Tricep Skinfold Thickness with Gestational Metabolic Profiles in White European and South Asian Women
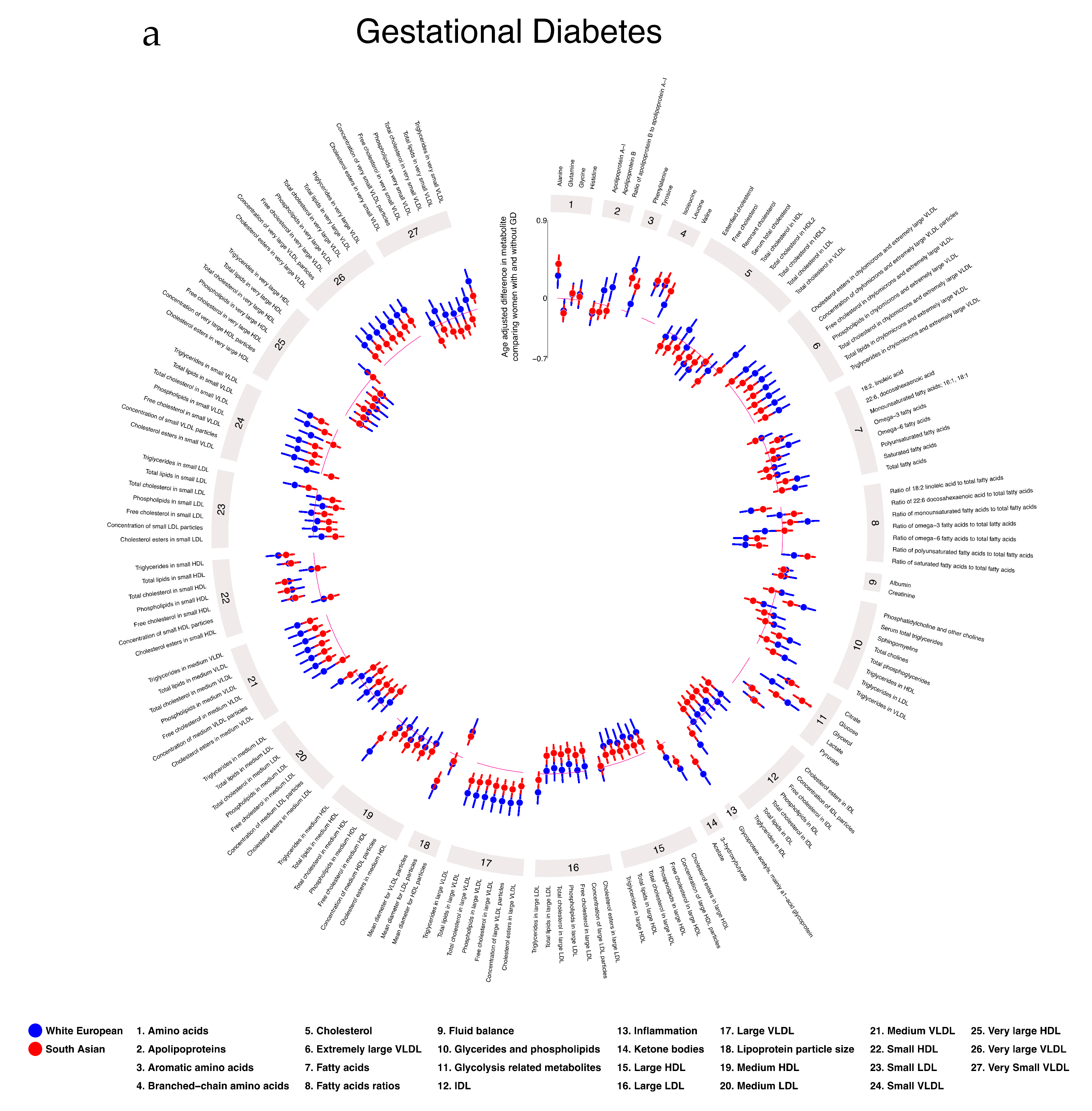
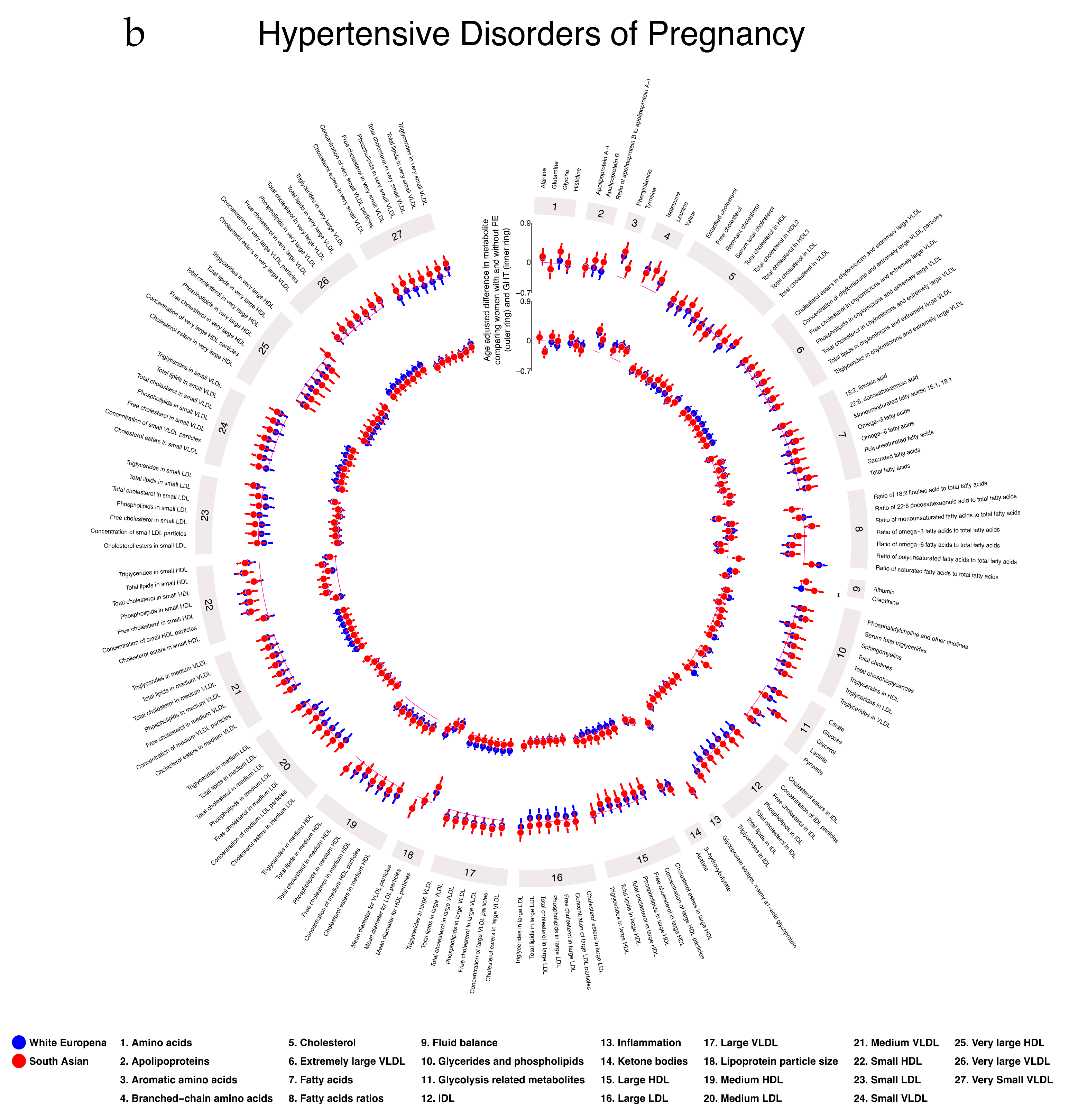
2.5. Associations of Gestational Diabetes, Pre-Eclampsia, and Gestational Hypertension with Gestational Metabolic Profiles in White European and South Asian Women
2.6. Additional Analyses
3. Discussion
4. Materials and Methods
4.1. Participants
4.2. Assessment of Ethnicity
4.3. Maternal Pregnancy Measurements
4.4. Maternal Pregnancy Metabolic Profiling using the NMR Platform
4.4.1. Sample Collection and Storage
4.4.2. NMR Protocol
4.4.3. Metabolite Quantification and Quality Control
4.4.4. Validation of the NMR Platform
4.5. Statistical Analyses
- Model 1: age-adjusted (except for association of age with metabolic profiles). Rationale—maternal age is known to influence metabolic profiles. Also, age can influence education, parity, height, BMI, TST, GD, GHT, and PE and, thus, could confound the association of any of these exposures with metabolic profiles.
- Model 2: for exposures height, BMI, TST, GD, GHT, and PE only, we adjusted for education and parity (in addition to age), as these could influence these exposures and metabolic profiles and, hence, could be confounders.
- Model 3: for GD, GHT, and PE only, we adjusted as for model 2 but additionally adjusted for BMI, as this might confound the association of these with metabolites.
Dealing with Missing Data
4.6. Additional Analyses
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Wang, Q.; Würtz, P.; Auro, K.; Mäkinen, V.-P.; Kangas, A.J.; Soininen, P.; Tiainen, M.; Tynkkynen, T.; Jokelainen, J.; Santalahti, K.; et al. Metabolic profiling of pregnancy: Cross-sectional and longitudinal evidence. BMC Med. 2016, 14, 205. [Google Scholar] [CrossRef] [PubMed]
- Mills, H.L.; Patel, N.; White, S.L.; Pasupathy, D.; Briley, A.L.; Santos Ferreira, D.L.; Seed, P.T.; Nelson, S.M.; Sattar, N.; Tilling, K.; et al. The effect of a lifestyle intervention in obese pregnant women on gestational metabolic profiles: Findings from the UK Pregnancies Better Eating and Activity Trial (UPBEAT) RCT. BMC Med. 2019, 17, 15. [Google Scholar] [CrossRef] [PubMed]
- Würtz, P.; Kangas, A.J.; Soininen, P.; Lawlor, D.A.; Smith, G.D.; Ala-Korpela, M. Quantitative Serum Nuclear Magnetic Resonance Metabolomics in Large-Scale Epidemiology: A Primer on Omic Technologies. Am. J. Epidemiol. 2017, 186, 1084–1096. [Google Scholar] [CrossRef] [PubMed]
- Auro, K.; Joensuu, A.; Fischer, K.; Kettunen, J.; Salo, P.; Mattsson, H.; Niironen, M.; Kaprio, J.; Eriksson, J.G.; Lehtimäki, T.; et al. A metabolic view on menopause and ageing. Nat. Commun. 2014, 5, 4708. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ferreira, D.L.S.; Williams, D.M.; Kangas, A.J.; Soininen, P.; Ala-Korpela, M.; Smith, G.D.; Järvelin, M.-R.; Lawlor, D.A. Association of pre-pregnancy body mass index with offspring metabolic profile: Analyses of 3 European prospective birth cohorts. PLoS Med. 2017, 14, e1002376. [Google Scholar]
- Würtz, P.; Wang, Q.; Kangas, A.J.; Richmond, R.C.; Skarp, J.; Tiainen, M.; Tynkkynen, T.; Soininen, P.; Havulinna, A.S.; Kaakinen, M.; et al. Metabolic Signatures of Adiposity in Young Adults: Mendelian Randomization Analysis and Effects of Weight Change. PLoS Med. 2014, 11, e1001765. [Google Scholar] [CrossRef]
- Bell, J.A.; Hamer, M.; Richmond, R.C.; Timpson, N.J.; Carslake, D.; Smith, G.D. Associations of device-measured physical activity across adolescence with metabolic traits: Prospective cohort study. PLoS Med. 2018, 15, e1002649. [Google Scholar] [CrossRef]
- Würtz, P.; Cook, S.; Wang, Q.; Tiainen, M.; Tynkkynen, T.; Kangas, A.J.; Soininen, P.; Laitinen, J.; Viikari, J.; Kähönen, M.; et al. Metabolic profiling of alcohol consumption in 9778 young adults. Int. J. Epidemiol. 2016, 45, 1493–1506. [Google Scholar] [CrossRef]
- Vogt, S.; Wahl, S.; Kettunen, J.; Breitner, S.; Kastenmüller, G.; Gieger, C.; Suhre, K.; Waldenberger, M.; Kratzsch, J.; Perola, M.; et al. Characterization of the metabolic profile associated with serum 25-hydroxyvitamin D: A cross-sectional analysis in population-based data. Int. J. Epidemiol. 2016, 45, 1469–1481. [Google Scholar] [CrossRef]
- Bell, J.A.; Carslake, D.; Wade, K.H.; Richmond, R.C.; Langdon, R.J.; Vincent, E.E.; Holmes, M.V.; Timpson, N.J.; Smith, G.D. Influence of puberty timing on adiposity and cardiometabolic traits: A Mendelian randomisation study. PLoS Med. 2018, 15, e1002641. [Google Scholar] [CrossRef]
- Wang, Q.; Würtz, P.; Auro, K.; Morin-Papunen, L.; Kangas, A.J.; Soininen, P.; Tiainen, M.; Tynkkynen, T.; Joensuu, A.; Havulinna, A.S.; et al. Effects of hormonal contraception on systemic metabolism: cross-sectional and longitudinal evidence. Int. J. Epidemiol. 2016, 45, 1445–1457. [Google Scholar] [CrossRef] [PubMed]
- Sliz, E.; Kettunen, J.; Holmes, M.V.; Williams, C.O.; Boachie, C.; Wang, Q.; Männikkö, M.; Sebert, S.; Walters, R.; Lin, K.; et al. Metabolomic Consequences of Genetic Inhibition of PCSK9 Compared with Statin Treatment. Circulation 2018, 138, 2499–2512. [Google Scholar] [CrossRef]
- Würtz, P.; Wang, Q.; Soininen, P.; Kangas, A.J.; Fatemifar, G.; Tynkkynen, T.; Tiainen, M.; Perola, M.; Tillin, T.; Hughes, A.D.; et al. Metabolomic Profiling of Statin Use and Genetic Inhibition of HMG-CoA Reductase. J. Am. Coll. Cardiol. 2016, 67, 1200–1210. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Desert, R.; Canlet, C.; Costet, N.; Cordier, S.; Bonvallot, N. Impact of maternal obesity on the metabolic profiles of pregnant women and their offspring at birth. Metabolomics 2015, 11, 1896–1907. [Google Scholar] [CrossRef] [Green Version]
- Diaz, S.O.; Pinto, J.I.M.; Graça, G.; Duarte, I.F.; Barros, A.S.; Galhano, E.; Pita, C.; Almeida, M.D.C.; Goodfellow, B.J.; Carreira, I.M.; et al. Metabolic Biomarkers of Prenatal Disorders: An Exploratory NMR Metabonomics Study of Second Trimester Maternal Urine and Blood Plasma. J. Proteome Res. 2011, 10, 3732–3742. [Google Scholar] [CrossRef]
- Dudzik, D.; Zórawski, M.; Skotnicki, M.; Zarzycki, W.; Kozlowska, G.; Bibik-Malinowska, K.; Vallejo, M.; Garcia, A.; Barbas, C.; Ramos, M.P.; et al. Metabolic fingerprint of Gestational Diabetes Mellitus. J. Proteom. 2014, 103, 57–71. [Google Scholar] [CrossRef]
- Pinto, J.I.M.; Almeida, L.M.; Martins, A.S.; Duarte, D.; Barros, A.S.; Galhano, E.; Pita, C.; Almeida, M.D.C.; Carreira, I.M.; Gil, A.M. Prediction of Gestational Diabetes through NMR Metabolomics of Maternal Blood. J. Proteome Res. 2015, 14, 2696–2706. [Google Scholar] [CrossRef]
- Sandler, V.; Reisetter, A.C.; Bain, J.R.; Muehlbauer, M.J.; Nodzenski, M.; Stevens, R.D.; Ilkayeva, O.; Lowe, L.P.; Metzger, B.E.; Newgard, C.B.; et al. Associations of maternal BMI and insulin resistance with the maternal metabolome and newborn outcomes. Diabetologia 2017, 60, 518–530. [Google Scholar] [CrossRef]
- Huynh, J.; Xiong, G.; Bentley-Lewis, R. A systematic review of metabolite profiling in gestational diabetes mellitus. Diabetologia 2014, 57, 2453–2464. [Google Scholar] [CrossRef] [Green Version]
- Bahado-Singh, R.; Poon, L.C.; Yilmaz, A.; Syngelaki, A.; Turkoglu, O.; Kumar, P.; Kirma, J.; Allos, M.; Accurti, V.; Li, J.; et al. Integrated Proteomic and Metabolomic prediction of Term Preeclampsia. Sci. Rep. 2017, 7, 16189. [Google Scholar] [CrossRef]
- Kelly, R.S.; Giorgio, R.T.; Chawes, B.L.; Palacios, N.I.; Gray, K.J.; Mirzakhani, H.; Wu, A.; Blighe, K.; Weiss, S.T.; Lasky-Su, J. Applications of Metabolomics in the Study and Management of Preeclampsia; A Review of the Literature. Metabolomics 2017, 13, 86. [Google Scholar] [CrossRef] [PubMed]
- Kuc, S.; Koster, M.P.H.; Pennings, J.L.A.; Hankemeier, T.; Berger, R.; Harms, A.C.; Dane, A.D.; Schielen, P.C.J.I.; Visser, G.H.A.; Vreeken, R.J. Metabolomics Profiling for Identification of Novel Potential Markers in Early Prediction of Preeclampsia. PLoS ONE 2014, 9, e98540. [Google Scholar] [CrossRef] [PubMed]
- De Seymour, J.V.; Conlon, C.A.; Sulek, K.; Bôas, S.G.V.; McCowan, L.M.E.; Kenny, L.C.; Baker, P.N. Early pregnancy metabolite profiling discovers a potential biomarker for the subsequent development of gestational diabetes mellitus. Acta Diabetol. 2014, 51, 887–890. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- He, X.; de Seymour, J.V.; Sulek, K.; Qi, H.; Zhang, H.; Han, T.L.; Villas-Bôas, S.G.; Baker, P.N. Maternal hair metabolome analysis identifies a potential marker of lipid peroxidation in gestational diabetes mellitus. Acta Diabetol. 2016, 53, 119–122. [Google Scholar] [CrossRef] [PubMed]
- White, S.L.; Pasupathy, D.; Sattar, N.; Nelson, S.M.; Lawlor, D.A.; Briley, A.L.; Seed, P.T.; Welsh, P.; Poston, L. Metabolic profiling of gestational diabetes in obese women during pregnancy. Diabetologia 2017, 60, 1903–1912. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yajnik, C.S.; Yudkin, J.S. The Y-Y paradox. Lancet 2004, 363, 163. [Google Scholar] [CrossRef]
- McKeigue, P.M.; Pierpoint, T.; Ferrie, J.E.; Marmot, M.G. Relationship of glucose intolerance and hyperinsulinaemia to body fat pattern in south Asians and Europeans. Diabetologia 1992, 35, 785–791. [Google Scholar] [PubMed]
- Sattar, N.; Gill, J.M.R. Type 2 diabetes in migrant south Asians: Mechanisms, mitigation, and management. Lancet Diabetes Endocrinol. 2015, 3, 1004–1016. [Google Scholar] [CrossRef]
- Whincup, P.H.; Nightingale, C.M.; Owen, C.G.; Rudnicka, A.R.; Gibb, I.; Mckay, C.M.; Donin, A.S.; Sattar, N.; Alberti, K.G.M.M.; Cook, D.G. Early emergence of ethnic differences in type 2 diabetes precursors in the UK: The child heart and health study in England (CHASE study). PLoS Med. 2010, 7, e1000263. [Google Scholar] [CrossRef] [PubMed]
- Nightingale, C.M.; Krishnaveni, G.V.; Rudnicka, A.R.; Owen, C.G.; Veena, S.R.; Hill, J.C.; Cook, D.G.; Fall, C.H.D.; Whincup, P.H. Cardiometabolic risk markers in Indian children: Comparison with UK Indian and White European children. PLoS ONE 2012, 7, e36236. [Google Scholar] [CrossRef] [PubMed]
- Yajnik, C.S.; Lubree, H.G.; Rege, S.S.; Naik, S.S.; Deshpande, J.A.; Deshpande, S.S.; Joglekar, C.V.; Yudkin, J.S. Adiposity and Hyperinsulinemia in Indians Are Present at Birth. J. Clin. Endocrinol. Metab. 2002, 87, 5575–5580. [Google Scholar] [CrossRef] [Green Version]
- West, J.; Lawlor, D.A.; Fairley, L.; Bhopal, R.; Cameron, N.; McKinney, P.A.; Sattar, N.; Wright, J. UK-born Pakistani-origin infants are relatively more adipose than white British infants: Findings from 8704 mother-offspring pairs in the Born-in-Bradford prospective birth cohort. J. Epidemiol. Community Health 2013, 67, 544–551. [Google Scholar] [CrossRef] [PubMed]
- West, J.; Wright, J.; Fairley, L.; Sattar, N.; Whincup, P.; Lawlor, D.A. Do ethnic differences in cord blood leptin levels differ by birthweight category? Findings from the born in bradford cohort study. Int. J. Epidemiol. 2014, 43, 249–254. [Google Scholar] [CrossRef] [PubMed]
- Farrar, D.; Fairley, L.; Santorelli, G.; Tuffnell, D.; Sheldon, T.A.; Wright, J.; Van Overveld, L.; Lawlor, D.A. Association between hyperglycaemia and adverse perinatal outcomes in south Asian and white British women: Analysis of data from the Born in Bradford cohort. Lancet Diabetes Endocrinol. 2015, 3, 795–804. [Google Scholar] [CrossRef]
- Wright, J.; Small, N.; Raynor, P.; Tuffnell, D.; Bhopal, R.; Cameron, N.; Fairley, L.; Lawlor, D.A.; Parslow, R.; Petherick, E.S.; et al. Cohort profile: The born in bradford multi-ethnic family cohort study. Int. J. Epidemiol. 2013, 42, 978–991. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Scholl, T.O.; Stein, T.P.; Steer, R.A.; Williams, K.P. Maternal Circulating Lipid Profile during Early Pregnancy: Racial/Ethnic Differences and Association with Spontaneous Preterm Delivery. Nutrients 2017, 9, 19. [Google Scholar] [CrossRef]
- Koukkou, E.; Watts, G.F.; Mazurkiewicz, J.; Lowy, C. Ethnic differences in lipid and lipoprotein metabolism in pregnant women of African and Caucasian origin. J. Clin. Pathol. 1994, 47, 1105–1107. [Google Scholar] [CrossRef]
- Schreuder, Y.J.; Hutten, B.A.; Van Eijsden, M.; Jansen, E.H.; Vissers, M.N.; Twickler, M.T.; Vrijkotte, T.G.M. Ethnic differences in maternal total cholesterol and triglyceride levels during pregnancy: The contribution of demographics, behavioural factors and clinical characteristics. Eur. J. Clin. Nutr. 2011, 65, 580–589. [Google Scholar] [CrossRef]
- Van Eijsden, M.; Hornstra, G.; Van Der Wal, M.F.; Bonsel, G.J. Ethnic differences in early pregnancy maternal n-3 and n-6 fatty acid concentrations: An explorative analysis. Br. J. Nutr. 2009, 101, 1761–1768. [Google Scholar] [CrossRef]
- Catov, J.M.; Ness, R.B.; Wellons, M.F.; Jacobs, D.R.; Roberts, J.M.; Gunderson, E.P. Prepregnancy Lipids Related to Preterm Birth Risk: The Coronary Artery Risk Development in Young Adults Study. J. Clin. Endocrinol. Metab. 2010, 95, 3711–3718. [Google Scholar] [CrossRef] [Green Version]
- Vrijkotte, T.G.M.; Krukziener, N.; Hutten, B.A.; Vollebregt, K.C.; Van Eijsden, M.; Twickler, M.B. Maternal Lipid Profile During Early Pregnancy and Pregnancy Complications and Outcomes: The ABCD Study. J. Clin. Endocrinol. Metab. 2012, 97, 3917–3925. [Google Scholar] [CrossRef] [PubMed]
- Smedts, H.; Van Uitert, E.; Valkenburg, O.; Laven, J.; Eijkemans, M.; Lindemans, J.; Steegers, E.; Steegers-Theunissen, R. A derangement of the maternal lipid profile is associated with an elevated risk of congenital heart disease in the offspring. Nutr. Metab. Cardiovasc. Dis. 2012, 22, 477–485. [Google Scholar] [CrossRef] [PubMed]
- Ritchie, S.C.; Würtz, P.; Nath, A.P.; Abraham, G.; Havulinna, A.S.; Fearnley, L.G.; Sarin, A.-P.; Kangas, A.J.; Soininen, P.; Aalto, K.; et al. The Biomarker GlycA Is Associated with Chronic Inflammation and Predicts Long-Term Risk of Severe Infection. Cell Syst. 2015, 1, 293–301. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Deelen, J.; Kettunen, J.; Fischer, K.; van der Spek, A.; Trompet, S.; Kastenmüller, G.; Boyd, A.; Zierer, J.; van den Akker, E.B.; Ala-Korpela, M.; et al. A metabolic profile of all-cause mortality risk identified in an observational study of 44,168 individuals. Nat. Commun. 2019, 10, 3346. [Google Scholar] [CrossRef] [PubMed]
- Wang, T.J.; Larson, M.G.; Vasan, R.S.; Cheng, S.; Rhee, E.P.; McCabe, E.; Lewis, G.D.; Fox, C.S.; Jacques, P.F.; Fernandez, C.; et al. Metabolite Profiles and the Risk of Developing Diabetes. Nat. Med. 2011, 17, 448–453. [Google Scholar] [CrossRef] [PubMed]
- Würtz, P.; Mäkinen, V.-P.; Soininen, P.; Kangas, A.J.; Tukiainen, T.; Kettunen, J.; Savolainen, M.J.; Tammelin, T.; Viikari, J.S.; Rönnemaa, T.; et al. Metabolic Signatures of Insulin Resistance in 7098 Young Adults. Diabetes 2012, 61, 1372–1380. [Google Scholar] [PubMed]
- Magnusson, M.; Lewis, G.D.; Ericson, U.; Orho-Melander, M.; Hedblad, B.; Engström, G.; Östling, G.; Clish, C.; Wang, T.J.; Gerszten, R.E.; et al. A diabetes-predictive amino acid score and future cardiovascular disease. Eur. Heart J. 2013, 34, 1982–1989. [Google Scholar] [CrossRef]
- Monrose, E.; Zamudio, A.R.; Aristide, A.; Navalurkar, R.; Bedekar, R.; Ferrara, L.; Kushner, T. Sa1562 – Intrahepatic Cholestasis of Pregnancy (ICP) is Associated with Higher Prevalence of Nafld: A Case-Control Study. Gastroenterology 2019, 156, 1235–1236. [Google Scholar] [CrossRef]
- Hartley, A.; Ferreira, D.L.S.; Anderson, E.L.; Lawlor, D.A. Metabolic profiling of adolescent non-alcoholic fatty liver disease. Wellcome Open Res. 2018, 3, 166. [Google Scholar] [CrossRef]
- Manta-Vogli, P.D.; Schulpis, K.H.; Dotsikas, Y.; Loukas, Y.L. The significant role of amino acids during pregnancy: Nutritional support. J. Matern. Neonatal Med. 2018, 28, 1–7. [Google Scholar] [CrossRef]
- Lawlor, D.A.; Tilling, K.; Smith, G.D. Triangulation in aetiological epidemiology. Int. J. Epidemiol. 2016, 45, 1866–1886. [Google Scholar] [CrossRef] [PubMed]
- Taylor, K.; Santos Ferreira, D.L.; Lawlor, D. Analysis Plan: Differences in Pregnancy Metabolic Profiles and Their Determinants between White European and South Asian Women: Findings from the Born in Bradford Cohort. Available online: https://osf.io/v7swp/ (accessed on 8th May 2019).
- Esko, T.; Hirschhorn, J.N.; Feldman, H.A.; Hsu, Y.-H.H.; Deik, A.A.; Clish, C.B.; Ebbeling, C.B.; Ludwig, D.S. Metabolomic profiles as reliable biomarkers of dietary composition. Am. J. Clin. Nutr. 2017, 105, 547–554. [Google Scholar] [CrossRef] [PubMed]
- Kujala, U.M.; Mäkinen, V.P.; Heinonen, I.; Soininen, P.; Kangas, A.J.; Leskinen, T.H.; Rahkila, P.; Würtz, P.; Kovanen, V.; Cheng, S.; et al. Long-term leisure-time physical activity and serum metabolome. Circulation 2013, 127, 340–348. [Google Scholar] [CrossRef] [PubMed]
- Lawlor, D.A.; West, J.; Fairley, L.; Nelson, S.M.; Bhopal, R.S.; Tuffnell, D.; Freeman, D.J.; Wright, J.; Whitelaw, D.C.; Sattar, N. Pregnancy glycaemia and cord-blood levels of insulin and leptin in Pakistani and white British mother-offspring pairs: Findings from a prospective pregnancy cohort. Diabetologia 2014, 57, 2492–2500. [Google Scholar] [CrossRef] [PubMed]
- Kannieappan, L.M.; Deussen, A.R.; Grivell, R.M.; Yelland, L.; Dodd, J.M. Developing a tool for obtaining maternal skinfold thickness measurements and assessing inter-observer variability among pregnant women who are overweight and obese. BMC Pregnancy Childbirth 2013, 13, 42. [Google Scholar] [CrossRef] [PubMed]
- Würtz, P.; Havulinna, A.S.; Soininen, P.; Tynkkynen, T.; Prieto-Merino, D.; Tillin, T.; Ghorbani, A.; Artati, A.; Wang, Q.; Tiainen, M.; et al. Metabolite profiling and cardiovascular event risk: A prospective study of 3 population-based cohorts. Circulation 2015, 131, 774–785. [Google Scholar] [CrossRef] [PubMed]
- Kettunen, J.; Tukiainen, T.; Sarin, A.-P.; Ortega-Alonso, A.; Tikkanen, E.; Lyytikäinen, L.-P.; Kangas, A.J.; Soininen, P.; Würtz, P.; Silander, K.; et al. Genome-wide association study identifies multiple loci influencing human serum metabolite levels. Nat. Genet. 2012, 44, 269–276. [Google Scholar] [CrossRef] [PubMed] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Maternal Characteristics | Category | All (n = 8774) | White European (n = 4072) | South Asian (n = 4702) | Diff in Means or OR (95% CI) * |
|---|---|---|---|---|---|
| Age, years | 27.3 ± 5.6 | 26.7 ± 6.0 | 27.8 ± 5.2 | 1.1 (0.8, 1.3) | |
| Height (cm) | 161.7 ± (6.4) | 164.2 ± 6.2 | 159.5 ± 5.8 | 4.7 (4.5, 5.0) | |
| Missing (%) | 172 (2.0) | 57 (1.4) | 115 (2.4) | - | |
| BMI (kg/m2) | 26.1 ± 5.7 | 26.7 (6.0) | 25.6 ± 5.4 | 1.1 (0.9, 1.4) | |
| Missing (%) | 413 (4.7) | 183 (4.5) | 230 (4.9) | - | |
| TST (mm) | 25.4 ± 7.1 | 25.7 ± 7.2 | 24.6 (6.9) | 1.1 (0.6, 1.6) | |
| Missing (%) a | 5671 (64.6) | 1891 (46.4) | 3780 (80.4) | - | |
| Education | Below A-level | 5151 (58.7) | 2462 (60.5) | 2689 (57.2) | Ref |
| A-level or above | 3446 (39.3) | 1523 (37.4) | 1923 (40.9) | 1.2 (1.1, 1.3) | |
| Unknown/Missing (%) | 177 (2.0) | 87 (2.1) | 90 (1.9) | - | |
| HDP | Normotensive | 7902 (90.1) | 3533 (86.8) | 4369 (92.9) | Ref |
| PE | 224 (2.6) | 118 (2.9) | 106 (2.3) | 0.7 (0.6, 0.9) | |
| GHT | 634 (7.2) | 417 (10.2) | 217 (4.6) | 0.4 (0.3, 0.5) | |
| Missing (%) | 14 (0.2) | 4 (0.1) | 10 (0.2) | - | |
| Gestational Diabetes | Yes | 734 (8.4) | 209 (5.1) | 525 (11.2) | 2.3 (1.9, 2.7) |
| Parity | Median (IQR) | 1 (0–2) | 1 (0–1) | 1 (0–2) | - |
| Nulliparous | 3433 (39.1) | 1938 (47.6) | 1495 (31.8) | Ref | |
| Multiparous | 5037 (57.4) | 2000 (49.1) | 3037 (64.6) | 2.0 (1.8, 2.1) | |
| Missing (%) | 304 (3.5) | 134 (3.3) | 170 (3.6) | - | |
| Gest Age at Blood Sampling (weeks) | 26.3 (2.0) | 26.2 (1.9) | 26.3 (2.0) | 0.0 (−0.1, 0.0) |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Taylor, K.; L. Santos Ferreira, D.; West, J.; Yang, T.; Caputo, M.; A. Lawlor, D. Differences in Pregnancy Metabolic Profiles and Their Determinants between White European and South Asian Women: Findings from the Born in Bradford Cohort. Metabolites 2019, 9, 190. https://doi.org/10.3390/metabo9090190
Taylor K, L. Santos Ferreira D, West J, Yang T, Caputo M, A. Lawlor D. Differences in Pregnancy Metabolic Profiles and Their Determinants between White European and South Asian Women: Findings from the Born in Bradford Cohort. Metabolites. 2019; 9(9):190. https://doi.org/10.3390/metabo9090190
Chicago/Turabian StyleTaylor, Kurt, Diana L. Santos Ferreira, Jane West, Tiffany Yang, Massimo Caputo, and Deborah A. Lawlor. 2019. "Differences in Pregnancy Metabolic Profiles and Their Determinants between White European and South Asian Women: Findings from the Born in Bradford Cohort" Metabolites 9, no. 9: 190. https://doi.org/10.3390/metabo9090190