Article Text

Abstract
BACKGROUND Glucagon-like peptide-1(7-36)amide (GLP-1) is a gut hormone released postprandially. Synthetic GLP-1 strongly inhibits gastric emptying in healthy subjects and in patients with diabetes mellitus.
AIMS To investigate the effects of GLP-1 on antro-pyloro-duodenal motility in humans.
METHODS Eleven healthy male volunteers were studied on two separate days. On the interdigestive study day, a basal period was followed by a 60 minute period of saline infusion and two further 60 minute periods of intravenous infusion of GLP-1 0.4 and 1.2 pmol/kg/min to achieve postprandial and supraphysiological plasma levels, respectively. On the postprandial study day, the same infusions were coadministered with intraduodenal lipid perfusion at 2.5 ml/min (2.5 kcal/min) followed by another 60 minutes of recording after cessation of GLP-1. Antro-pyloro-duodenal motility was measured by perfusion manometry.
RESULTS GLP-1 significantly inhibited the number and amplitudes of antral and duodenal contractions in the interdigestive state and after administration of duodenal lipid. It abolished interdigestive antral wave propagation. In the interdigestive state, GLP-1 dose dependently increased pyloric tone and significantly stimulated isolated pyloric pressure waves (IPPW). Pyloric tone increased with duodenal lipid, and this was further enhanced by GLP-1. GLP-1 transiently restored the initial IPPW response to duodenal lipid which had declined with lipid perfusion. Plasma levels of pancreatic polypeptide were dose dependently diminished by GLP-1 with and without duodenal lipid.
CONCLUSIONS GLP-1 inhibited antro-duodenal contractility and stimulated the tonic and phasic motility of the pylorus. These effects probably mediate delayed gastric emptying. Inhibition of efferent vagal activity may be an important mechanism. As postprandial plasma levels of GLP-1 are sufficient to appreciably affect motility, we believe that endogenous GLP-1 is a physiological regulator of motor activity in the antro-pyloro-duodenal region.
- GLP-1
- gastrointestinal motility
- pylorus
- pancreatic polypeptide
Statistics from Altmetric.com
The proglucagon derived glucagon-like peptide-1(7-36)amide (GLP-1) is a gastrointestinal hormone released postprandially from the L cells of the gut.1 2 It exerts a glucose dependent insulinotropic effect at the pancreatic B cell and lowers glucagon release.3-5 GLP-1 may also enhance glucose uptake in peripheral tissues6 and has recently been shown to mediate satiety in humans.7 8 These combined effects improve glucose tolerance and are the rationale for evaluating its therapeutic potential in the treatment of diabetes mellitus.9-11
Previous studies in humans have shown that synthetic GLP-1 substantially retards gastric emptying of liquid and solid meals.11-13 In addition to its insulinotropic and glucagonostatic action, reduction of the gastric emptying rate may be involved in the glucose lowering effect of GLP-1 in healthy subjects and in patients with diabetes mellitus.11 13 Independent of the tonic pressure generated by the proximal stomach, transpyloric pulsatile flow regulated by the motility of the antro-pyloro-duodenal region is a major mechanism in gastric emptying.14 15Antral and especially antro-duodenal coordinated contractions were shown to be associated with the gastric emptying rate of liquids2 16 17 and solids.18 Tonic and localised increases in phasic pressure generated by the pylorus provide an important braking mechanism diminishing gastric outflow.15 19-21
In this study, we have evaluated the effects of graded doses of synthetic GLP-1 on the motility of the antro-pyloro-duodenal region during the interdigestive and postprandial states in humans, the latter being elicited by duodenal perfusion of lipid. Duodenal perfusion of lipid was used instead of oral ingestion of a meal to provide a constant duodenal nutrient load independent of gastric emptying. This particular meal was chosen to establish a stable postprandial motility pattern and to minimise changes in plasma glucose and insulin. The latter are considerable under exogenous GLP-1 and concomitant hyperglycaemia. Finally, we compared the effects of GLP-1 in relation to lipids which is the classic physiological stimulator of pyloric motility.
Material and methods
SUBJECTS
Eleven healthy male volunteers, 23–28 years old and within 10% of ideal body weight, participated in the studies. None was receiving medications and none suffered from gastrointestinal symptoms or any systemic disease. The studies were approved by the ethics committee of the medical faculty of the Philipps-University of Marburg, and all participants gave written informed consent.
MOTILITY RECORDING
Perfusion manometry was recorded using a nine lumen duodenal sleeve/side hole catheter (Dentsleeve, South Australia, Australia). The manometric assembly incorporated a 4.5 cm long sleeve sensor, two antral side holes (2 cm apart), and three duodenal side holes (2 cm apart), beginning at the proximal and distal end of the sleeve, respectively. Two further side holes were positioned across the sleeve 1.5 cm apart. An additional lumen located 12 cm distal to the sleeve sensor was used for duodenal perfusion.
The correct position of the duodenal probe with the sleeve array straddling the pylorus was fluoroscopically checked before the start of each experiment and monitored throughout each study by measuring the transmucosal potential difference at the distal antral and the proximal duodenal port, as previously described.2 A difference of at least −15 mV indicated the correct transpyloric position of the tube.
The motility channels were perfused at a rate of 0.3 ml/min using a low compliance pneumohydraulic pump (Arndorfer Medical Specialists, Greendale, Wisconsin). Pressures were measured by external transducers. Data were simultaneously recorded on the screen of a multichannel chart system (PC Polygraph, Synectics Medical, Stockholm, Sweden) and stored in the memory of a personal computer. Data were sampled and digitised at 8 Hz followed by digital smoothing by a factor of 2.
EXPERIMENTAL PROTOCOL
All studies were performed after an overnight fast. Two experiments separated by at least one week were performed on each subject in random order with subjects lying comfortably in a semirecumbent position. An indwelling catheter was inserted into an antecubital vein for intravenous infusions. A second catheter was inserted in a retrograde manner into a dorsal vein of the contralateral hand for sampling of arterialised venous blood.13
Each experiment was started after a basal period of at least 30 minutes, the last 15 minutes of which displayed low motor activity (<5 antral contractions/10 minutes). On the interdigestive study day, saline 0.154 M was continuously perfused into the lower duodenal flexure at a rate of 2.5 ml/min throughout the experiment. In the interdigestive experiments, a 60 minute period of intravenous infusion of saline was followed by two further 60 minute periods of intravenous infusion of GLP-1 0.4 and 1.2 pmol/kg/min, to achieve physiological postprandial and supraphysiological plasma levels, respectively. During the initial 10 minutes of each infusion period, GLP-1 was infused at twice the dose (0.8 and 2.4 pmol/kg/min, respectively) to rapidly establish steady state plasma levels. On the postprandial study day, the same intravenous infusions were administered together with intraduodenal lipid perfusion at 2.5 kcal/min (2.5 ml/min, Lipofundin MCT 10%, Braun Co., Melsungen, Germany). This lipid preparation consisted of 50% medium chain triglycerides and 50% long chain triglycerides (soya bean oil). In the postprandial experiments, measurements were continued for another 60 minutes with lipid perfusion after cessation of intravenous GLP-1 to determine recovery characteristics. Antro-pyloro-duodenal motility was continuously recorded throughout all studies.
Blood samples were taken at −10 minutes and immediately before the start of intravenous infusion in addition to 10 minute intervals thereafter throughout each experiment. Blood was collected in ice chilled EDTA tubes containing aprotinin 1000 kallikrein inhibitory units/ml blood and centrifuged immediately. Plasma was stored at −20°C until assayed.
ANALYSIS OF MOTILITY TRACINGS
Motility recordings were analysed only when the transmucosal potential difference indicated the correct transpyloric position of the tube. Analysis of contractile events was computer assisted, applying validated software.22 Only peaks with amplitudes of at least 10 mm Hg and durations of at least 2 s were considered true contractions. Duodenal phase III was defined as the occurrence of regular contractions at a frequency ⩾10/minute for at least 2 minutes in the duodenum propagated aborally.
Data were analysed in 10 minute segments for the antrum and duodenum separately by summarising (frequency, motility index) or averaging (amplitude) the values derived from the two antral and three duodenal side holes, respectively. Motility indexes were determined as area under the contractions and expressed in mm Hg•s/min. Antral contractions were judged to be antro-pyloro-duodenally propagated waves if the onset of the pressure wave recorded in the most proximal duodenal side hole occurred within five seconds after the onset of a pressure wave recorded in one of the antral side holes, and if they were registered by all side holes in between.
Two parameters were used to describe pyloric motility: (1) isolated pyloric pressure waves (IPPW) were defined as contractions registered by the sleeve with or without simultaneous contractions in one sleeve side hole in the absence of associated (±three seconds) waves of any amplitude in the adjacent antral and duodenal side holes; (2) pyloric tone was measured by subtracting the basal pressure recorded by the antral side hole at the proximal end of the sleeve from the basal pressure recorded by the sleeve. Basal pressures (i.e. mean pressures after excluding contractions) were obtained for each minute and were used to calculate mean pyloric tone.
GLP-1(7-36)AMIDE
Synthetic GLP-1(7-36)amide was purchased from Wherl Co. (Wolfenbüttel, Germany). It has a peptide content of 87.1% and a peptide purity >99%. It was dissolved in 1% human serum albumin, filtered through 0.2 μm nitrocellulose filters and stored at −70°C.13 Samples were tested for pyrogens and bacterial growth, and no contamination with bacteria or endotoxins was detected. Dose calculations were based on the peptide content of the preparation.
DETERMINATIONS AND ASSAYS
Plasma glucose concentrations were measured by a glucose analyser using the glucose oxidase method (YSI 1500 G; Schlag, Bergisch-Gladbach, Germany) with a coefficient of variation of <2%. Plasma insulin was measured using the Abbott Imx Microparticle Enzyme Immunoassay, with an average intra-assay coefficient of variation of 5%. Plasma immunoreactivities of C peptide, glucagon, and pancreatic polypeptide (PP) were analysed using commercially available radioimmunoassay kits (Biermann, Bad Nauheim, Germany and Euradiagnostica, the Netherlands (PP)). Immunoreactive GLP-1 was measured using the specific polyclonal antibody GA 1178 (Affinity Research, Nottingham, UK), as described previously.2 The detection limit of the assay was 0.25 pmol/l. The antiserum did not cross react with glucose dependent insulinotropic peptide (GIP), pancreatic glucagon, glicentin, oxyntomodulin, or GLP-2. Intra- and interassay coefficients of variation were 3.4% and 10.4%, respectively.
STATISTICAL ANALYSIS
All values were expressed as mean (SEM). Plasma and motility parameters were analysed separately for each 60 minute period. Pyloric tone was calculated as change from basal, the latter being determined as mean pyloric tone during the total basal period before the start of the experiments. Differences in plasma hormones and glucose compared with the basal state were calculated as integrated values over basal (area under the response curve; AUC). Basal values for plasma parameters were determined as the mean of the two basal values just before the start of each experiment. All samples were tested for normality using the Komolgorov-Smirnov test. Differences between experimental sets for each parameter were analysed by two way repeated measures analysis of variance using intravenous infusion and duodenal perfusion as factors. When this analysis indicated differences, a Student-Newman-Keuls multicomparison test was performed. Differences were considered significant at p<0.05.
Results
Intravenous infusion of GLP-1 dose dependently increased plasma GLP-1 with both duodenal saline and lipid reaching constant plasma levels within 20 minutes after the start of infusion (fig 1, table 1). Steady state plasma levels were 6.2 (0.6) pmol/l (low dose GLP-1) and 13.1 (0.9) pmol/l (high dose GLP-1) with duodenal saline, and 7.1 (0.6) pmol/l (low dose GLP-1) and 14.4 (0.9) pmol/l (high dose GLP-1) with duodenal lipid, respectively. Duodenal lipid slightly but significantly increased plasma GLP-1 compared with duodenal saline perfusion (2.4 (0.3) pmol/l v 1.4 (0.2) pmol/l; p<0.05). This difference remained constant with infusion of both loads of exogenous GLP-1. After cessation of GLP-1 during lipid perfusion, plasma levels of GLP-1 immediately decreased and gradually returned to preinfusion values.
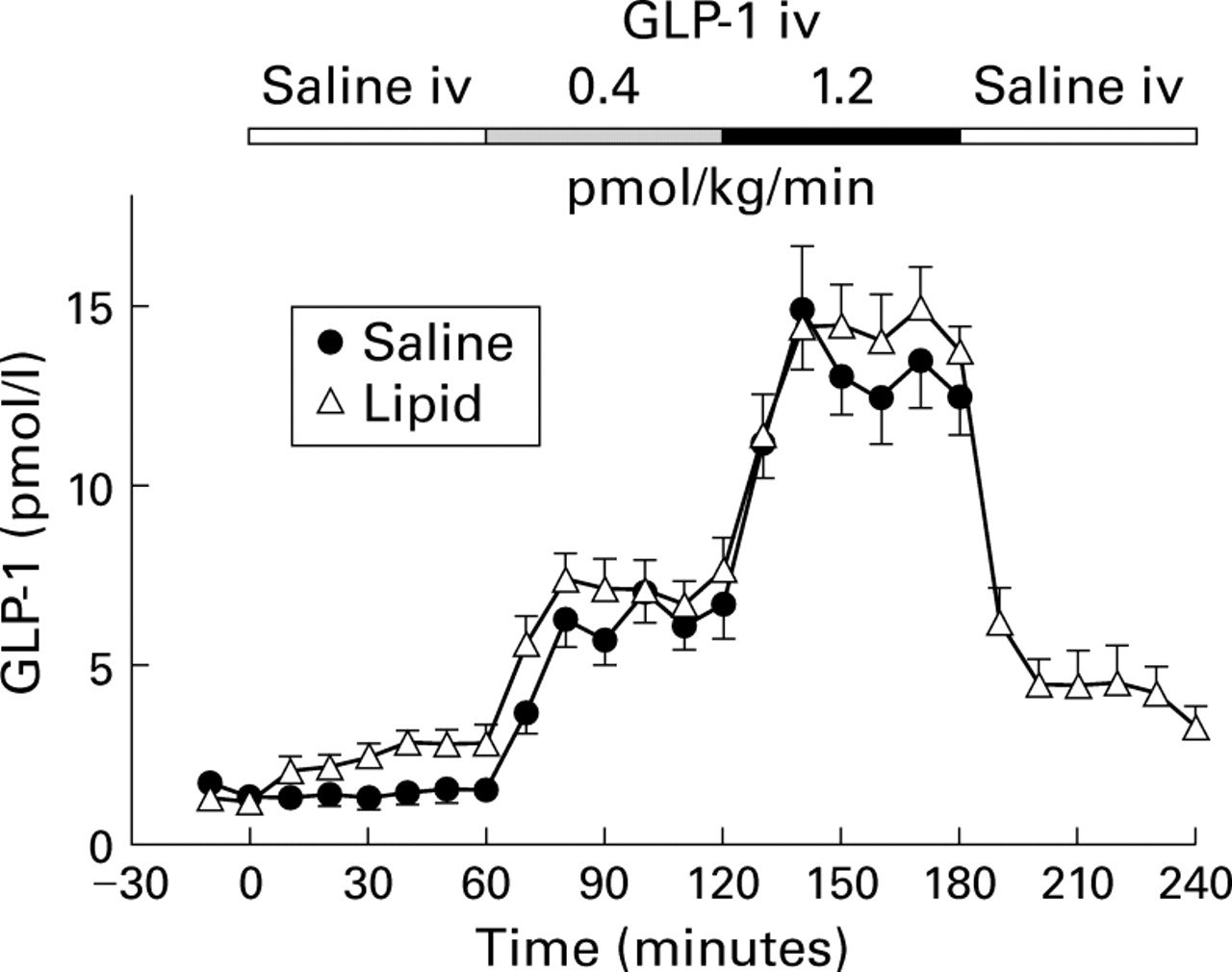
Plasma immunoreactivities of GLP-1 in response to intravenous infusions of saline, and GLP-1(7-36)amide 0.4 and 1.2 pmol/kg/min with concomitant duodenal perfusion of saline or lipid 2.5 kcal/min in 11 healthy volunteers (mean (SEM)). For statistical analysis, see table 1.
Effect of physiological (low dose, 0.4 pmol/kg/min) and supraphysiological (high dose, 1.2 pmol/kg/min) doses of GLP-1 on plasma glucose, and plasma immunoreactivities of GLP-1, insulin, glucagon, and pancreatic polypeptide with intraduodenal perfusion of saline or lipid (2.5 kcal/min, 2.5 ml/min)
ANTRO-DUODENAL AND PYLORIC MOTILITY
The transmucosal potential difference indicated the correct transpyloric position of the probe during 95.9 (1.6)% and 92.5 (2.0)% of recording time with duodenal saline and lipid, respectively.
In the interdigestive experiments with duodenal saline perfusion, both doses of GLP-1 significantly inhibited the number, amplitude, and motility indexes of contractions in the antrum and duodenum (fig 2A, table 2). Inhibition of antro-duodenal motility instantaneously (within 2 minutes) occurred after the start of the low dose infusion of GLP-1. Within 20 minutes, even the low dose of GLP-1 almost completely inhibited antro-duodenal motility. Antral waves propagating across the pylorus to the duodenum over a distance of 4.5 cm and 6.5 cm, respectively, were abolished, even by the low dose of GLP-1 (table 3).

Contraction frequencies (top) and amplitudes (bottom) in the antrum and duodenum in response to intravenous infusions of saline, and GLP-1(7-36)amide 0.4 and 1.2 pmol/kg/min during concomitant duodenal perfusion of saline (A) or lipid 2.5 kcal/min (B) in 11 healthy volunteers (mean (SEM)). For statistical analysis, see table 2.
Effect of physiological (low dose, 0.4 pmol/kg/min) and supraphysiological (high dose, 1.2 pmol/kg/min) doses of GLP-1 on antral and duodenal motility with intraduodenal perfusion of saline or lipid (2.5 kcal/min, 2.5 ml/min)
Effect of physiological (low dose, 0.4 pmol/kg/min) and supraphysiological (high dose, 1.2 pmol/kg/min) doses of GLP-1 on pyloric motility and antro-pyloro-duodenal wave propagation with intraduodenal perfusion of saline or lipid (2.5 kcal/min, 2.5 ml/min)
Compared with duodenal saline, duodenal perfusion of lipid inhibited antral and duodenal contractility. Against a background of duodenal lipid, exogenous GLP-1 dose dependently diminished the number, amplitude, and motility indexes of antral and duodenal contractions to a comparable residual level as in the interdigestive state (fig 2B, table 2). With cessation of GLP-1, antro-duodenal motility immediately increased and returned to preinfusion activity. Duration of contractions remained unchanged with GLP-1 in all experiments (data not shown).
Pyloric tone dose dependently increased with exogenous GLP-1 in the interdigestive state (fig 3B, table 3). Even with the low dose of GLP-1, the effect was significant within 10 minutes and was fully established within 20 minutes after the start of infusion of GLP-1. With duodenal lipid, pyloric tone increased compared with duodenal saline, and this was enhanced further by GLP-1 (fig 3B, table 3). Pyloric tone steadily decreased after cessation of GLP-1 in parallel with declining plasma levels. During the last 30 minutes, pyloric tone was not significantly different from the first 60 minutes without GLP-1, indicating complete recovery (3.9 (0.7) mm Hg for the last 30 minutes v 3.1 (0.4) mm Hg during the first 60 minutes of the experiments; p=0.29).

Isolated pyloric pressure waves (IPPW) (A) and pyloric tone (B) in response to intravenous infusions of saline, and GLP-1(7-36)amide 0.4 and 1.2 pmol/kg/min during concomitant duodenal perfusion of saline (top) or lipid 2.5 kcal/min (bottom) in 11 healthy volunteers (mean (SEM)). *p<0.05 for comparison of times indicated by the arrows (paired t test). For further statistical analysis, see table3.
IPPWs immediately and significantly increased with infusion of GLP-1 (fig 3A, table 3). But in contrast with the tonic pyloric response, increases in IPPWs were not dose dependent and exhibited a fast increase followed by a pronounced decline. The latter phenomenon held true for both lipid alone and GLP-1 against a background of duodenal lipid. Stimulation of IPPW by duodenal lipid was significantly stronger than with GLP-1 alone, and duodenal lipid and exogenous GLP-1 did not act in an additive manner. With cessation of GLP-1, IPPWs further decreased and were significantly diminished compared with the initial 60 minutes without GLP-1.
In the interdigestive state with both doses of GLP-1, initiation of phasic and tonic stimulation of the pylorus was almost paralleled by duodenal phase III-like episodes, occurring within 10 minutes after the start of GLP-1 (fig 4). These duodenal phase III-like episodes were demonstrated in seven of 11 volunteers after the low dose and in five subjects in response to the high dose of GLP-1. With intraduodenal lipid, seven of 11 volunteers immediately developed a duodenal phase III in parallel with pyloric stimulation. The typical pattern of the antro-pyloro-duodenal motility seen with GLP-1 infusion is shown in fig5.
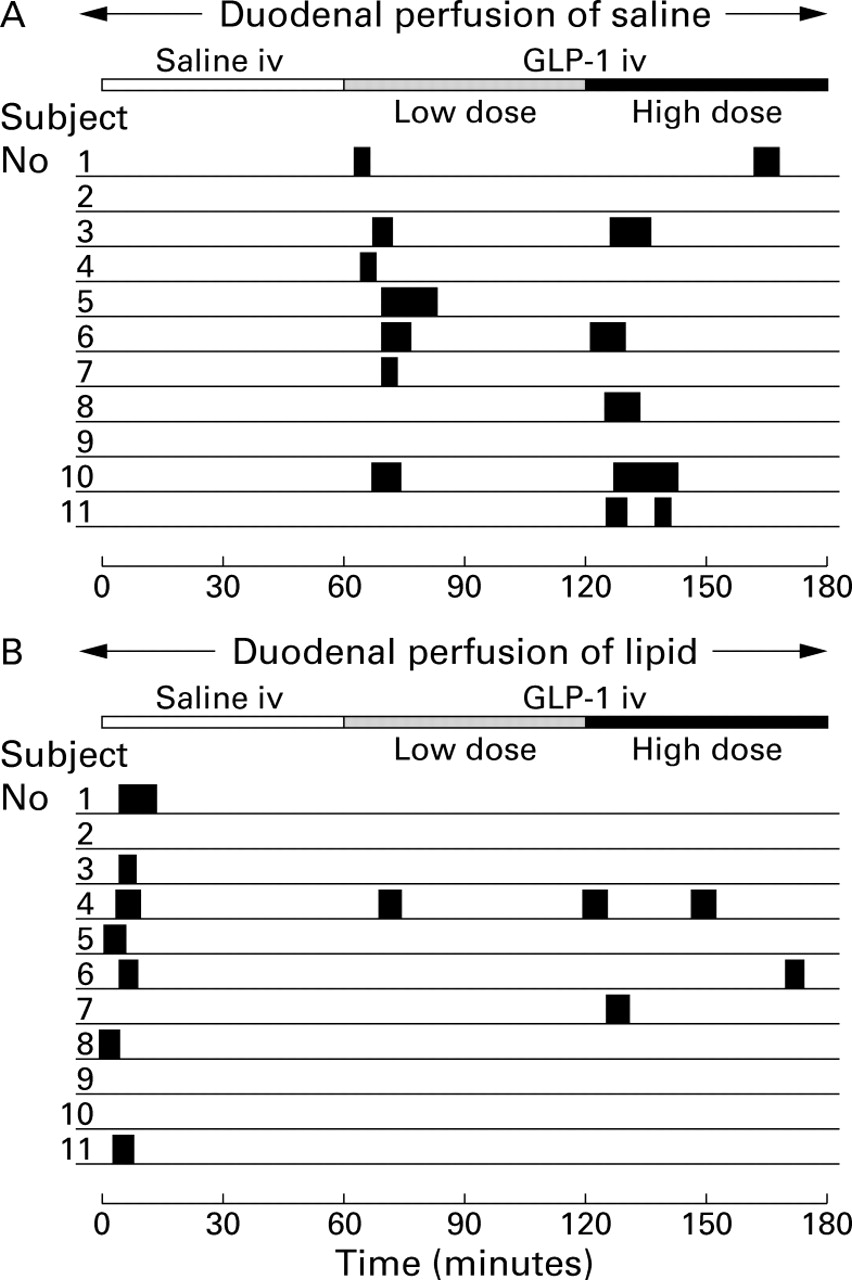
Occurrence of duodenal phase III-like activity with duodenal perfusion of saline (A) or lipid 2.5 kcal/min (B) in 11 healthy volunteers. In seven of 11 subjects, an activity front was seen within 10 minutes after the start of the low dose infusion of GLP-1 in the interdigestive state or duodenal lipid perfusion in the postprandial studies, respectively. The length of the solid bars represents the length of contraction burst.

Manometric tracings showing the effects of intravenous infusion of GLP-1 0.4 pmol/kg/min during duodenal perfusion of saline (A) and lipid 2.5 kcal/min (B). In the interdigestive state (A), GLP-1 immediately inhibited antro-duodenal motility and induced a sustained increase in basal pyloric pressure with concomitant short lasting stimulation of IPPWs. During duodenal lipid perfusion (B), antral and duodenal contractility were completely abolished by GLP-1, and basal pyloric pressure further increased in addition to the effect of lipid alone, paralleled by stimulation of IPPWs.
PLASMA GLUCOSE AND HORMONES
In the interdigestive experiments, basal plasma glucose was 4.3 (0.1) mmol/l and significantly decreased with both doses of GLP-1 (fig6A, table 1). With duodenal lipid, plasma glucose slightly decreased compared with duodenal saline perfusion. This decrease was further pronounced with both doses of GLP-1 followed by a significant increase after cessation of GLP-1. However, the nadir of plasma glucose was 3.58 (0.12) mmol/l and 3.64 (0.14) mmol/l at 20 minutes with the low and high doses of GLP-1, respectively. These nadir values did not represent hypoglycaemia.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Effects of intravenous infusions of saline, and GLP-1(7-36)amide 0.4 and 1.2 pmol/kg/min on plasma glucose (A) and immunoreactivities of glucagon (B), insulin (C), and pancreatic polypeptide (D) during concomitant duodenal perfusion of saline or lipid 2.5 kcal/min in 11 healthy volunteers. Mean (SEM) of incremental values over basal. For statistical analysis, see table 1.
Mean basal plasma levels of insulin were 6.0 (0.7) mU/l and 6.1 (0.7) mU/l before the start of duodenal saline and lipid, respectively. Both doses of GLP-1 caused a short initial increase in insulin followed by a decline (fig 6C). Compared with duodenal saline, duodenal lipid elicited a small but significant increase in plasma insulin which was maintained throughout each study (table 1).
Basal levels of glucagon were 91.5 (8.8) pg/ml and 88.2 (8.6) pg/ml in the studies with duodenal saline and lipid, respectively (NS). With duodenal saline and lipid, GLP-1 significantly and dose dependently diminished plasma glucagon (fig 6B) followed by an immediate increase with cessation of GLP-1 in the lipid experiments. Compared with saline perfusion, duodenal lipid significantly increased glucagon levels which remained significantly elevated throughout the experiments (table 1).
Basal levels of pancreatic polypeptide (PP) averaged 64.3 (8.6) pg/ml and 71.9 (8.7) pg/ml before the start of duodenal saline and lipid, respectively (p=0.32). With duodenal lipid compared with saline perfusion, levels of PP increased greatly (fig 6D, table 1). Intravenous GLP-1 dose dependently diminished PP with and without duodenal lipid, and in the experiments with duodenal saline, PP was significantly reduced below basal levels. After cessation of GLP-1, an instantaneous increase in pancreatic polypeptide levels was observed in the studies with duodenal lipid perfusion, indicating complete recovery.
Discussion
Our study is the first to address the effects of GLP-1 on the motility of the antro-pyloro-duodenal region in humans. Physiological plasma levels of GLP-1 were reached with low doses of synthetic GLP-1. They corresponded to peak levels found after oral ingestion of a high caloric glucose meal.2 23 With the high dose of GLP-1, supraphysiological plasma levels were obtained.
In the interdigestive state and with duodenal lipid perfusion, GLP-1 greatly inhibited antro-duodenal contractility by reduction of contraction frequencies and amplitudes. It abolished antral wave propagation in the interdigestive state. The tonic and phasic motility of the pylorus was stimulated by GLP-1 with and without duodenal lipid. GLP-1 dose dependently and reversibly inhibited pancreatic polypeptide release, indicating inhibition of efferent vagal-cholinergic activity. All effects were obtained at postprandial plasma levels of GLP-1.
Previous studies in humans have shown that synthetic GLP-1 substantially retards gastric emptying of liquid and solid meals in a dose dependent manner.11-13 A major mechanism of gastric emptying is pulsatile transpyloric flow which is mediated by gastric and propagated antro-pyloric phasic contractions.14 15 In humans, antral and especially antro-pyloro-duodenally propagated contractions are associated with the gastric emptying rate of liquids2 16 17 and solids.18 Tonic and phasic increases in pressure generated by the pylorus provide an important braking mechanism diminishing the stroke volume of gastric outflow.15 19-21 Therefore, we suggest that the following motor effects of GLP-1 are important mediators of impaired gastric emptying: (i) inhibition of antral waves in general, (ii) reduction of transpyloric propagated antral waves in particular, and (iii) simultaneous stimulation of localised phasic and tonic pyloric contractions.
In dogs, a similar motor pattern with infusion of synthetic GLP-1 has been demonstrated to be associated with reduction in transpyloric flow and inhibition of gastric emptying of a non-caloric liquid meal.24 In humans, subcutaneously injected GLP-1 dose dependently inhibited antral and coordinated antroduodenal contractions in parallel with prolongation of the lag phase of gastric emptying of a mixed liquid meal.13 In addition, preliminary data obtained from human studies indicate strong relaxation of the proximal stomach in response to intravenous GLP-1 using the same doses as in the present study.25 Thus exogenous GLP-1 reduces driving forces and stimulates braking mechanisms of gastric outflow thus impacting on all motor sites known to determine gastric emptying.
The effects on antroduodenal and pyloric motility appeared at physiological postprandial plasma levels of GLP-1. Therefore, endogenous GLP-1 may be a physiological regulator of motor activity in the antro-pyloro-duodenal region. The postprandial release of GLP-1 depends on the gastric emptying rate of nutrients. A temporal association between duodenal delivery of glucose, retardation of gastric emptying, and an increase in plasma GLP-1 has been demonstrated previously.2 Preliminary studies using the specific GLP-1 antagonist exendin(9-39)amide revealed that endogenous GLP-1 inhibited the intestinal phase of antro-duodenal motility during duodenal glucose perfusion in humans.26 Gastric emptying of nutrient containing meals is regulated by intestinal feedback mechanisms triggered by small intestinal chemoreceptors.27-29 In accordance with a previous study,30 duodenal lipid perfusion elicited a postprandial motor pattern with inhibition of antro-duodenal motility and strong stimulation of pyloric tone and IPPWs. As endogenous GLP-1 is significantly released with duodenal lipid, it may at least partially mediate the motility response to duodenal lipid. Pyloric motility is primarily under neural control,31 32 but humoral signals may modulate the pyloric response.33 34 Cholecystokinin (CCK), which is also released with duodenal nutrients, stimulates pyloric motility similar to GLP-1.35 Therefore, endogenous CCK may also account in part for pyloric motility stimulated by duodenal lipid. But in contrast with GLP-1, endogenous CCK has been shown to stimulate duodenal motility in the intestinal phase34 36; thus the motility effects of both peptides do not seem to be entirely complementary. However, the effects of endogenous GLP-1 on gastrointestinal motility remain to be established.
Comparison of pyloric stimulation by GLP-1 with and without duodenal lipid provides new insights in the regulation of pyloric motility. Pyloric tone increased steadily with both duodenal lipid and exogenous GLP-1 against a background of duodenal saline, the latter in a dose dependent manner. The tonic response to duodenal lipid was enhanced further by exogenous GLP-1 in parallel with increasing GLP-1 plasma levels. GLP-1 stimulated pyloric tone at least as strongly as duodenal lipid, and the interaction was additive. In contrast, increases in IPPWs with step doses of GLP-1 were not dose dependent, and GLP-1, even at a supraphysiological dose, was unable to stimulate IPPWs as strongly as duodenal lipid, despite much higher plasma levels. Thus the phasic pyloric response to duodenal lipid may depend less on GLP-1 release, and the mechanisms whereby GLP-1 and lipid stimulate pyloric motility may be different, at least in terms of IPPWs.
Irrespective of the mode of stimulation, the IPPW response was attenuated after an initial increase, in contrast with pyloric tone. With duodenal lipid, GLP-1 transiently restored the IPPW response which declined with lipid perfusion. After cessation of GLP-1, the frequencies of IPPW greatly decreased below the level of the first 60 minutes, despite constant lipid perfusion, whereas tone remained significantly elevated. In contrast with the sustained but variable tonic response, the phasic pyloric response to different stimuli showed a stereotype pattern with fast attenuation. This suggests that the tonic pyloric response is of greater physiological importance. In dogs, dose dependent retardation of gastric emptying by graded doses of intravenous GLP-1 was accompanied by a dose dependent increase in pyloric tone but not of IPPWs.24 Furthermore, the different sensitivities of the tonic and phasic pyloric responses to atropine,37 to enzymatic blockade of nitric oxide synthase,38 and to hyperglycaemia39 may indicate different regulation mechanisms of these two motor phenomena of the pylorus.
In most of the volunteers, GLP-1 rapidly triggered duodenal phase III-like episodes in the interdigestive state. Duodenal phase III-like activity has been demonstrated previously in response to a variety of stimuli, such as intraduodenal infusion of dextrose20 and lipid,30 intravenous β endorphin,40 during stress induced by cold,41 pain,42 or labyrinthine stimulation,43 and acute hyperglycaemia.39 Interestingly, intraduodenal lipid stimulated duodenal phase III-like episodes in the present study. However, with lipid perfusion, the start of infusion of GLP-1 was accompanied by duodenal phase III-like episodes in only two of 11 subjects. All duodenal phase III-like episodes strongly paralleled initiation of pyloric stimulation, either by GLP-1 or duodenal lipid. Therefore, this initial phasic and transient stimulation of the duodenum may originate in the pylorus and, although its functional significance is still uncertain, it may indicate an unspecific reaction in response to strong stimulation of the pyloric muscle by GLP-1 or lipid.
Inhibition of pancreatic polypeptide (PP), a hormone of the endocrine pancreas under strong vagal cholinergic control, was also found in humans administered subcutaneous or intravenous GLP-1 after oral ingestion of a meal.11 13 44 However, intestinal stimulation of PP release requires stimulation of enteropancreatic cholinergic reflexes by duodenal delivery of nutrients,45and a reduced duodenal nutrient load during retarded gastric emptying would explain this PP response. In the present study, PP release triggered by lipid perfusion directly into the duodenum was dose dependently diminished by GLP-1 followed by rapid and complete recovery after cessation of GLP-1. Moreover, in the interdigestive studies, PP was significantly reduced below basal levels. Therefore, we suggest that GLP-1 inhibits efferent vagal-cholinergic activity, thereby diminishing PP release and at least contributes to inhibition of antral and duodenal motility via a central pathway. Receptors for GLP-1 are present in circumventricular organs such as the subfornical organ, the nucleus of the solitary tract and the area postrema.46 In addition, it has been shown recently in rats that GLP-1 induced inhibition of gastric emptying involves a capsaicin sensitive pathway, indicating an interaction with vagal afferent nerves.47 A direct action of GLP-1 on pancreatic PP cells or a paracrine effect via somatostatin seems unlikely as GLP-1 induces stimulation rather than inhibition of PP release from isolated human pancreatic islets.48
However, inhibition of vagal-cholinergic input does not explain stimulation of the pylorus by GLP-1 because the pylorus is stimulated and not inhibited by a cholinergic pathway in humans.34 37 The existence of cholinergic excitatory neurones within the duodenal wall and non-adrenergic non-cholinergic innervation with nitric oxide mediating pyloric relaxation have been demonstrated in dogs.31 32 Here, we speculate that GLP-1 interacts with neurones of the enteric nervous system projecting to the pylorus.
This study was designed to minimise changes in plasma glucose and insulin with GLP-1, preventing interaction with gastrointestinal motility. Decreases in glucose were small and outside the hypoglycaemic range. Also, changes in plasma insulin were weak and within the fasting range. The insulinotropic activity of GLP-1 is highly glucose dependent but during euglycaemia, a small amount of insulin was released, as in previous studies.3 49 In contrast with insulin, inhibition of glucagon release by GLP-1 is not glucose dependent, and even basal endogenous GLP-1 has been shown to tonically inhibit glucagon release from the pancreatic A cell in humans during eu- and hyperglycaemia.5 Both inhibition of glucagon and stimulation of insulin may account for the diminution in plasma glucose in the present study.
The increase in insulin with duodenal lipid alone was intriguing, and was also accompanied by a weak decrease in plasma glucose. However, a small increase in insulin was observed in studies with oral50 and jejunal51 application of fat. Duodenal lipid caused a rapid increase in endogenous GLP-1, achieving plasma levels similar to those after oral ingestion of 50 g of glucose.2 Therefore, GLP-1 released by duodenal lipid may contribute to its weak insulinotropic effect. This is in agreement with a marked increase in insulin if fat is administered enterally during hyperglycaemia.50-52
Acute hyperglycaemia has been shown to inhibit antral motility53 and to stimulate phasic but not tonic pyloric motility in humans.39 Also, glucagon is a potent inhibitor of gastrointestinal motility.54 In this study, plasma glucose and glucagon decreased with GLP-1 infusion and thus they do not mediate the inhibitory motor effects of GLP-1. Euglycaemic hyperinsulinaemia with plasma insulin, increasing at least 10-fold over basal, has been shown to inhibit interdigestive antral and duodenal as well as postprandial jejunal motility in normal subjects.51 55 However, the slight increase in plasma insulin presented here is unlikely to account for the pronounced motility effects of duodenal lipid or synthetic GLP-1.
In conclusion, GLP-1 inhibited antro-duodenal and stimulated pyloric motility in the interdigestive and postprandial states in humans. This arguably contributes to inhibition of gastric emptying by GLP-1. Inhibition of efferent vagal-cholinergic activity may be involved in the inhibitory action of GLP-1.
DIGESTIVE DISORDERS FOUNDATION—CURRENT RESEARCH FELLOWSHIPS
Dr R Aspinall. Imperial College of Science, Technology & Medicine Research. Genetic vaccination for pancreatic cancer. Training fellowship. Awarded £37 361 (first year); £40 117 (estimated) (second year). Started October 1998. Duration two years. Sponsored by Amelie Waring.
Dr A Bertoletti. University College London. Dynamics of the virus specific immune response in hepatitis B virus infection. Research grant. Awarded £103 499 (first year); £73 156 (second year). Started September 1999. Duration two years. Sponsored by the National Lottery Charities Board.
Miss R Bright-Thomas. University College London Medical School. A murine clinical trial of gene therapy in familial adenomatous polyposis. Research training fellowship. Awarded £34 983 plus consumables. Started October 1999. Duration one year. Sponsored by AP&T/DDF.
Dr MJ Carter. University of Sheffield. The interleukin-1 gene cluster in ulcerative colitis: a family based association study. Research training fellowship. Awarded £33 931 plus consumables (first year); £36 302 (est.) plus consumables (second year). Started March 1999. Duration two years. Sponsored by ia (The Ileostomy and Internal Pouch Support Group).
Dr R Cunliffe. University of Nottingham. Isolation of novel antimicrobial peptides expressed by human small intestinal paneth cells. Research training fellowship. Awarded £25 196 (first year); £26 414 (second year). Started February 1998. Duration two years. Sponsored by APT VI.
Dr D Hobday. University of Manchester. The physiology of colorectal symptoms in health and in irritable bowel syndrome. Research training fellowship. Awarded £27 776 (first year); £29 255 (second year). Started February 1998. Duration two years. Sponsored by BDF.
Dr R Jones. University of Birmingham. The generation of tumour specific cytolytic T cells from human hepatocellular carcinoma. Medical research training fellowship. Awarded £29 775 (first year); £33 520 (second year); £36 762 (third year); plus tuition fees for years one to three. Started September 1997. Duration three years. Sponsored by BSG.
Dr A King. St Thomas's Hospital, London. A family linkage study to investigate the genetics of coeliac disease. Research training fellowship. Awarded £40 264 plus consumables (first year); £42 060 (est.) plus consumables (second year). Started November 1999. Duration two years. Sponsored by BSG/DDF.
Dr PA Kitchen. Northwick Park and St Mark's NHS Trust. Glucagon-like peptide 2 in the short bowel syndrome. Research training fellowship. Awarded £33 977 plus consumables. Started May 1999. Duration one year. Sponsored by AP&T/DDF.
Dr Y Mahida. University of Nottingham. Investigation of the role of bacteria in diarrhoeal disease. Research grant. Awarded £142 639 (first year); £111 220 (second year); £115 528 (third year). Started January 2000. Duration three years. Sponsored by the National Lottery Charities Board.
Miss K Martin. University of Manchester. Identification of mechanisms underlying age alterations in gut epithelium. Research training fellowship. Awarded £21 360 plus consumables (first year); £22 214 (est.) plus consumables (second year). Started July 1999. Duration two years. Sponsored by BDF/RIA.
Dr J McLaughlin. University of Manchester. Enteroendocrine cells: at the crossroads of abnormal brain-gut and gut-brain signalling across the gut epithelium. Senior clinical research fellowship. Awarded £50 000 per annum. Due to start Summer/ Autumn 2000. Duration four years. Sponsored by DDF.
Professor T Northfield. St George's Hospital Medical School. Helicobacter pylori infection: a treatable link between poverty and coronary heart disease? Research grant. Awarded £84 792 (first year); £113 525 (second year). Started February 1998. Duration two years. Sponsored by the National Lottery Charities Board.
Dr F Shek. University of Southampton. Mechanisms of stellate cell activation, proliferation and apoptosis in chronic pancreatitis. Research training fellowship. Awarded £31 638 (est.) plus consumables (first year); £33 294 (est.) plus consumables (second year). Started January 2000. Duration two years. Sponsored by Amelie Waring.
Mr D Slade. University of Manchester. The gastrointestinal tract in injury: villain or victim? Research training fellowship. Awarded £35 763 plus consumables (first year); £38 278 (est.) plus consumables (second year). Started September 1998. Duration two years. Sponsored by BDF/NRF.
Dr S Weaver. University of Bath. Colonic epithelial cell B lymphocyte interaction and subsequent COX II and iNOS expression. Research training fellowship. Awarded £26 135 (est.) plus consumables (first year); £27 199 (est.) plus consumables (second year). Started June 1999. Duration two years. Sponsored by DDF/NACC.
Acknowledgments
The excellent technical assistance of Gabriele Kraft and Michaela Junck is gratefully appreciated. We thank Mrs Elisabeth Bothe and Mrs Elke Birkenstock for measurement of plasma hormones and Mrs Maiszies for sterile preparation of GLP-1. This work was supported by a grant from the Deutsche Forschungsgemeinschaft.
References
Footnotes
- Abbreviations used in this paper:
- GLP-1
- glucagon-like peptide-1
- IPPW
- isolated pyloric pressure wave
- PP
- pancreatic polypeptide
- CCK
- cholecystokinin
- GIP
- glucose dependent insulinotropic peptide
Linked Articles
- Commentary