Article Text

Abstract
Introduction This study aims to determine whether high glucose condition and dynamic O-linked N-acetylglucosamine (O-GlcNAc) modification can promote the proliferation and migration of human retinal microvascular endothelial cells (HRMECs) and whether Runt-related transcription factor 1 (RUNX1) could mediate the glucose and O-GlcNAc-driven proliferation and migration of HRMECs.
Research design and methods Western blot analysis was used to detect the O-GlcNAc modification level and RUNX1 level in cells and retina tissues, cell growth was studied by cell counting kit-8 assay, cell proliferation was detected by immunofluorescence staining. Then, cell migration and tube formation were investigated by scratch-wound assay, Transwell assay, and tube-forming assay. The changes of retinal structure were detected by H&E staining. The O-GlcNAc modification of RUNX1 was detected by immunoprecipitation.
Results High glucose increases pan-cellular O-GlcNAc modification and the proliferation and migration of HRMECs. Hence, O-GlcNAc modification is critical for the proliferation and migration of HRMECs. RUNX1 mediates the glucose and O-GlcNAc-driven proliferation and migration in HRMECs. RUNX1 can be modified by O-GlcNAc, and that the modification is enhanced in a high glucose environment.
Conclusions The present study reveals that high glucose condition directly affects retinal endothelial cells (EC) function, and O-GlcNAc modification is critical for the proliferation and migration of HRMECs, RUNX1 may take part in this mechanism, and maybe the function of RUNX1 is related to its O-GlcNAc modification level, which provides a new perspective for studying the mechanism of RUNX1 in diabetic retinopathy.
- diabetes complications
Data availability statement
All data relevant to the study are included in the article.
This is an open access article distributed in accordance with the Creative Commons Attribution Non Commercial (CC BY-NC 4.0) license, which permits others to distribute, remix, adapt, build upon this work non-commercially, and license their derivative works on different terms, provided the original work is properly cited, appropriate credit is given, any changes made indicated, and the use is non-commercial. See: http://creativecommons.org/licenses/by-nc/4.0/.
Statistics from Altmetric.com
Significance of this study
What is already known about this subject?
High glucose can improve the level of O-linked N-acetylglucosamine (O-GlcNAc) modification of retina vascular endothelial cells (EC).
High glucose regulates Runt-related transcription factor 1 (RUNX1) and implicates RUNX1 in aberrant retinal angiogenesis.
What are the new findings?
High glucose condition directly affects retinal EC function.
O-GlcNAc modification is critical for the proliferation and migration of human retinal microvascular endothelial cells (HRMECs).
O-GlcNAc can modify RUNX1, and the modification is enhanced in a high glucose environment, and maybe the function of RUNX1 is related to its O-GlcNAc modification level.
How might these results change the focus of research or clinical practice?
Controlling the increase of O-GlcNAc modification may be beneficial for the occurrence and development of diabetic retinopathy (DR).
This research provides a new direction to study the mechanism of RUNX1 in DR; that is, we can explore the role of RUNX1 from the perspective of protein post-translational modification.
Introduction
Diabetic retinopathy (DR) is one of the complications caused by diabetes, and is the leading cause of blindness of the working-age population.1 DR can be divided into two stages: non-proliferative diabetic retinopathy and proliferative diabetic retinopathy (PDR). Neovascularization is a pathological feature of PDR.2 In recent days, a meta-analysis study that included 22 896 patients with diabetes revealed that for patients with diabetes, the prevalence of patients with DR was 34.6%. Furthermore, among these patients, the prevalence of PDR was 6.96%, while the prevalence of diabetic macular edema was 6.81%.3 The pathological changes of DR is retinal capillary endothelial damage, which includes selective loss of retinal capillary pericytes, a thickened basement membrane, occluded capillaries and vascular leakage, and these are due to the blood-retinal barrier damage. These changes lead to a wide range of retinal ischemia and retinal hypoxia, which eventually lead to the formation of new blood vessels. DR is a chronic disease. On the basis of hyperglycemia, multiple factors can lead to the occurrence and development of DR,4 5 such as the increased polyol pathway activity, the activation of the protein kinase C and hexosamine biosynthesis pathway (HBP), oxidative damage to the retina, and increased formation of advanced glycated end-products. However, the specific molecular mechanism remains not completely clear.
O-linked N-acetylglucosamine (O-GlcNAc) modification is a unique form of post-translational protein modification (PTM). Its uniqueness is embodied in its highly dynamic and revisible form of PTM. This can be regulated by two enzymes: O-GlcNAc transferase (OGT) and O-GlcNAcase (OGA). Furthermore, this uses UDP-GlcNAc as a donor, under the catalysis of OGT, it connects the O-glycosidic bond to protein serine/threonine residues, and under the catalysis of OGA, it hydrolyzes O-GlcNAc from serine/threonine residues.6 The changes in blood glucose levels can influence the UDP-GlcNAc level through the HBP pathway, and affect the O-GlcNAc modification of proteins. The most important feature of diabetes is sustained hyperglycemia. Elevated blood glucose can lead to increased UDP-GlcNAc donor, and subsequently lead to increased O-GlcNAc modification of proteins. Previous studies have revealed that O-GlcNAc modification plays an important role in diabetic cardiomyopathy and diabetic nephropathy.7 8 Recent studies have revealed that O-GlcNAc modification has similar effects in the process of DR.9 In high glucose (HG)-cultured retina endothelial cells, glycosyl transferases participate in the glycosylation process, and these glycosylation products change.10 In diabetic rat retina tissues, the O-GlcNAc modification of proteins increase.11
Neovascularization is a sign of PDR. Retinal neovascularization rupture leads to vitreous hemorrhage, and causes retinal vasoconstriction and traction retinal detachment, which are the main causes of visual impairment in patients with DR.12 13 Neovascularization is a dynamic and balanced process, which is regulated by angiogenic factors, such as vascular endothelial growth factor (VEGF) and antiangiogenic factors, such as pigment epithelium-derived factors (PEDFs).14 Under the action of angiogenic factors, vascular endothelial cells proliferate, migrate and form the lumen, and subsequently form new blood vessels. At present, the treatment for DR is neovascularization with the intraocular injection of anti-VEGF drugs, such as lucentis, ziv-aflibercept and conbercept, which have achieved good therapeutic effects. However, there are some side effects with the intraocular injection of anti-VEGF drugs, such as intraocular hemorrhage, endophthalmitis, neuroretinal injury, and ischemia-reperfusion injury.15–18 Therefore, ophthalmologists are still searching for new targets and the mechanism of DR neovascularization. Previous studies have found that in high-glucose cultured retinal cells, the O-GlcNAc modification of transcription factor specificity protein 1 (Sp1) can promote the expression of VEGF.19 The upregulation of VEGF expression is the direct cause of the neovascularization of DR.20 These above findings suggest that there are some relationships between O-GlcNAc modification and the formation of retinal neovascularization.
Runt-related transcription factor 1 (RUNX1) is a member of the Runx transcription factor family, and plays an important role in the determination of cell lineage differentiation direction, the formation of normal hematopoietic cells, and stem cell proliferation.21 RUNX1 was first obtained by the Miyoshi research group in 1991 in the leukemia cell clone of patients with acute myeloid leukemia. Hence, this was named AML1.22 Previous studies have revealed that the inhibition of RUNX1 can significantly inhibit the formation of retinal neovascularization in hypoxia-induced retinal neovascularization model mice. The knockdown of RUNX1 in human retinal microvascular endothelial cells (HRMECs) can reduce the ability of cells to form tubes,23 indicating that RUNX1 has a clear role in promoting angiogenesis. However, its mechanism remains unknown.
This present study investigates whether high glucose conditions and increased O-GlcNAc modification can promote the proliferation and migration of HRMECs and whether RUNX1 takes part in this process.
Materials and methods
Animals
Adult male Sprague-Dawley (SD) rats (250–300 g) were obtained from the Shanghai Laboratory Animal Center (Shanghai, China). Healthy male SD rats were randomly divided into two groups: wild-type (WT) group and diabetes mellitus group (DM).
Streptozotocin solution preparation method
The streptozotocin (STZ) powder was dissolved in 0.1 mol/L of sodium citrate buffer, and a 10 mg/mL STZ solution was obtained. Then, 0.22 µm of bacteria filter was used for filtration and sterilization.
The STZ (Sigma-Aldrich, St. Louis, USA) was injected into the rat’s abdomen (45 mg/kg). Then, the diabetes was confirmed by assaying the glucose concentration in blood, which was collected from the tail vein using a precision glucometer (Roche Diabetes Care, Mannheim, Germany) weekly after the STZ injection. Rats with a blood glucose concentration of >16.7 mmol/L were considered diabetic. All rats were sacrificed at 6 months after STZ injection.
Cells and treatments
The HRMECs were purchased from Cell Systems (Danvers, USA). The endothelial cell medium (ECM-2) was purchased from ScienCell (Carlsbad, USA), 500 mL medium contains 5% fetal bovine serum (FBS) (v/v), 1% penicillin and streptomycin (v/v), and 1% endothelial cell growth supplement (v/v).
Cells were treated with a medium containing 5.5 mM of glucose (normal (N)) or 25 mM of glucose (HG), and incubated at 37°C with 5% CO2. The OGA inhibitor Thiamet-G was purchased from Sigma-Aldrich, and the concentration used for the present experiments was 50 µM. The OGT inhibitor ST045849 was purchased from TimTec (Newark, USA), and the concentration used for the present experiments was 25 µM.
Immunoprecipitation
The SD rat retinas and cells were lysed using western blot analysis and immunoprecipitation (IP) lysis buffer (Sangon, Shanghai, China). Then, the protein concentration was assessed by bicinchoninic acid (BCA) assay. An appropriate amount of primary antibody was added to 1000 µg total protein (anti-O-GlcNAc antibody, PTM BioLabs (4 µg), anti-RUNX1 antibody from Santa Cruz (4 µg) and normal mouse IgG antibody from Cell Signaling Technology (4 µg)). The antigen-antibody complex was slowly shaken on the rotating shaker at 4°C overnight. Then, 35 µL of protein A/G agarose beads (Invitrogen, Carlsbad, USA) was added to the antigen-antibody complex, and slowly shaken on a rotating shaker at 4°C for 2 hours. Afterwards, the complex was washed for three times, the liquid was discarded, ddH2O and 2× protein loading was added to the sample, and this was heated at 100°C for 10 min. Then, the supernatant was collected and western blot analysis was performed.
Western blot analysis
The SD rat retinas and cells were lysed using western blot analysis and IP lysis buffer (Sangon, Shanghai, China). Then, the protein concentration was assessed by BCA assay. Afterwards, the samples were separated on 7.5% sodium dodecyl-sulfate polyacrylamide gel electrophoresis (SDS-PAGE) (separating gel: 2.5 mL acrylamide, 2.5 mL solution B, 0.1 mL 1% SDS (w/v), 4.8 mL ddH2O, 0.1 mL 1% ammonium persulfate (APS) (w/v), 10 µL Temed, stacking gel: 650 µL acrylamide, 1.25 mL solution D, 50 µL 1% SDS (w/v), 3 mL ddH2O, 50 µL 1% APS (w/v), 5 µL Temed, solution B and solution D were purchased from Songon Biotech, China). Then, the blots were transferred onto polyvinylidene fluoride membranes at 100 V for 65 min, blocked with 5% skimmed milk (w/v), and incubated with specific primary antibody at 4°C overnight. The antibodies used were anti-O-GlcNAc (1:1000), anti-RUNX1 (1:1000), and anti-β-actin (1:1000, Sangon, Shanghai, China).
Transfection
The transfections were performed according to instructions of RNAiMAX (Invitrogen, USA). The RUNX1 small interfering RNA (siRNA) was purchased from GenePharma (Shanghai, China).
Cell counting kit-8 assay
Cell counting kit-8 (CCK-8) assay was performed to detect cell proliferation. Different groups of HRMECs were plated at 96-well plate with 100 μL medium per well. After culture for 0, 24, 48 and 72 hours, 110 μL medium containing 10 µL CCK-8 (Yeasen, China) was added in each well. Cells were cultured for another 4 hours in incubator and then the absorbance was measured at a wavelength of 450 nm. Cell viability=[Ae−Ab]/[Ac−Ab], where Ae is absorbance of wells with cells, CCK-8 solution, and experimental reagents; Ab is absorbance of wells with medium and CCK-8 solution but without cells and Ac is absorbance of wells with cells and CCK-8 solution but without experimental reagents.
Immunofluorescence staining
Cells were fixed in 4% paraformaldehyde (w/v) for 20 min, permeabilized with 0.25% Triton X-100 (v/v) for 20 min, blocked in 2% BSA (w/v) for 20 min, and incubated in anti-ki67 (Cell Signaling Technology, Danvers, USA) overnight at a concentration of 1:400. Then, the liquid was discarded, the cells were incubated with the fluorescent secondary antibody (Invitrogen, Carlsbad, USA) at a concentration of 1:200. Cell counting was performed by ImageJ.
Scratch-wound migration assay
Different groups of HRMECs were plated in a 24-well plate with 500 µL of serum-free medium per well. Then, one scratch was generated per well, and the wound area was measured by ImageJ software at present and after 12 hours.
Transwell migration assay
The cell migration was accessed using a Transwell filter (Becton, Dickinson and Company). The cell suspension containing 1×105 cells was seeded on the upper chamber that contained 200 µL of serum-free medium, while 600 µL of ECM medium which contains 20% FBS (v/v) was added to the lower chamber. After culturing for 24 hours, these cells were fixed using 4% paraformaldehyde (w/v) for 20 min, and stained via 5% crystal violet solution (w/v) (Songon Biotech, China) for an additional 15 min. Cell counting was performed by ImageJ.
Tube-forming assay
Different groups of HRMECs were plated on a 96-well plate precoated with the basement membrane extract (Trevigen, USA), 100 µL of serum-free medium was add per well. Then, the cells were imaged at 24 hours after plating, and the tube formation was quantified using the Angiogenesis Analyzer plugin of ImageJ.
H&E staining
The rat eyeball was fixed with 4% paraformaldehyde (w/v), dehydrated by gradient alcohol, embedded in paraffin, and cut into a series of 5µm-thick slices. First, a slice was dewaxed by xylene, dehydrated with gradient ethanol, and stained with hematoxylin for 6 min, followed by running water wash for 10 min. After differentiation by 1% hydrochloric acid alcohol (v/v) for 15 s, the slice was rinsed by a trickle of water and counterstained with eosin for 3 min. On dehydration with gradient ethanol and transparency by xylene, the slice was sealed in neutral gum. The morphological changes in the retina were observed under an optical microscope.
Statistical analysis
The software used for statistical analysis is GraphPad prism 7. Statistical significance was determined using one-way analysis of variance. A p value <0.05 was considered statistically significant.
Results
Result 1: increased glucose concentration and O-GlcNAc modification is sufficient for proliferation and migration of HRMECs
The HRMECs were exposed to N or HG conditions, with or without Thiamet G. First, we found that 50 µM of Thiamet G could significantly increase the O-GlcNAc modification level in HRMECs. HG condition could also increase the O-GlcNAc modification level in HRMECs (figure 1A). Then we measured cell growth by CCK-8 assay. As shown in figure 1B, compared with group N, HG condition could promote cell growth. Treatment of cells with Thiamet G at the same glucose concentration could also promote cell growth. We used immunofluorescence staining to investigate the effects of HG condition and Thiamet G on cell proliferation. The trend of Ki67-positive rate was consistent with the trend of cell viability between different groups (figure 1C), this means that HG condition and high level of O-GlcNAc modification can promote cell proliferation. Scratch-wound migration assay and Transwell migration assay were selected to detect the migration ability of each group. Before that, we excluded the effect of apoptosis on migration detection, there was no change in apoptosis among the groups (figure 1D). Compared with group N, the wound closure rate increased by 16% and the total number of cells passing through the membrane nearly doubled under the condition of HG. At the same glucose concentration, treatment with Thiamet G could further increase the closure rate and the total number of cells passing through the membrane, this suggests that HG condition and high level of O-GlcNAc modification can promote cell migration (figure 1E,F). In addition, we detected the difference of tube formation in each group. We found that HG condition and high level of O-GlcNAc modification can also promote the formation of tube (figure 1G).

Increased glucose concentration and O-linked N-acetylglucosamine (O-GlcNAc) modification is sufficient for proliferation of human retinal microvascular endothelial cells (HRMECs). Cells were treated with 5.5 mM of glucose (normal (N)) or 5.5 mM+50 µM of Thiamet G (N+Thiamet G) or 25 mM of glucose (high glucose (HG)) or 25 mM of glucose+50 µM of Thiamet G (HG+Thiamet G). (A) The protein lysates were collected and processed by western blot analysis. Western blot analysis was performed using antibodies specific for O-GlcNAc modification and β-actin (a housekeeping gene). (B) Cell growth was determined by cell counting kit-8 (CCK-8), the graphs present the analysis of cell viability. (C) Cell proliferation was determined by Ki67 staining, the graphs present the analysis of ki67-positive rate. Scale bar=50 µm. (D) The apoptosis was detected by western blot analysis. Western blot analysis was performed using antibodies specific for cleaved-caspase 3, Bax, and β-actin. (E) Scratch-wound assay using HRMECs at 0 (top) and 12 hours (bottom). The black area represents the scratch area. The graphs represent the scratch healing rate. Scale bar=100 µm. (F) The migratory ability in HRMECs were detected by Transwell assay. The purple dots represent the cells that completed the vertical migration (passed through the membrane). The graphs represent the numbers of cells that passed through the membrane. Scale bar=100 µm. (G) Tube formation ability was detected by tube-forming assay. Cells were re-incubated on Matrigel. The graphs represent total segment length; n=3, independent experiments. Scale bar=100 µm. *P<0.05, **p<0.01, ***p<0.001, ****p<0.0001.
Result 2: O-GlcNAc modification is critical for the proliferation and migration of HRMECs
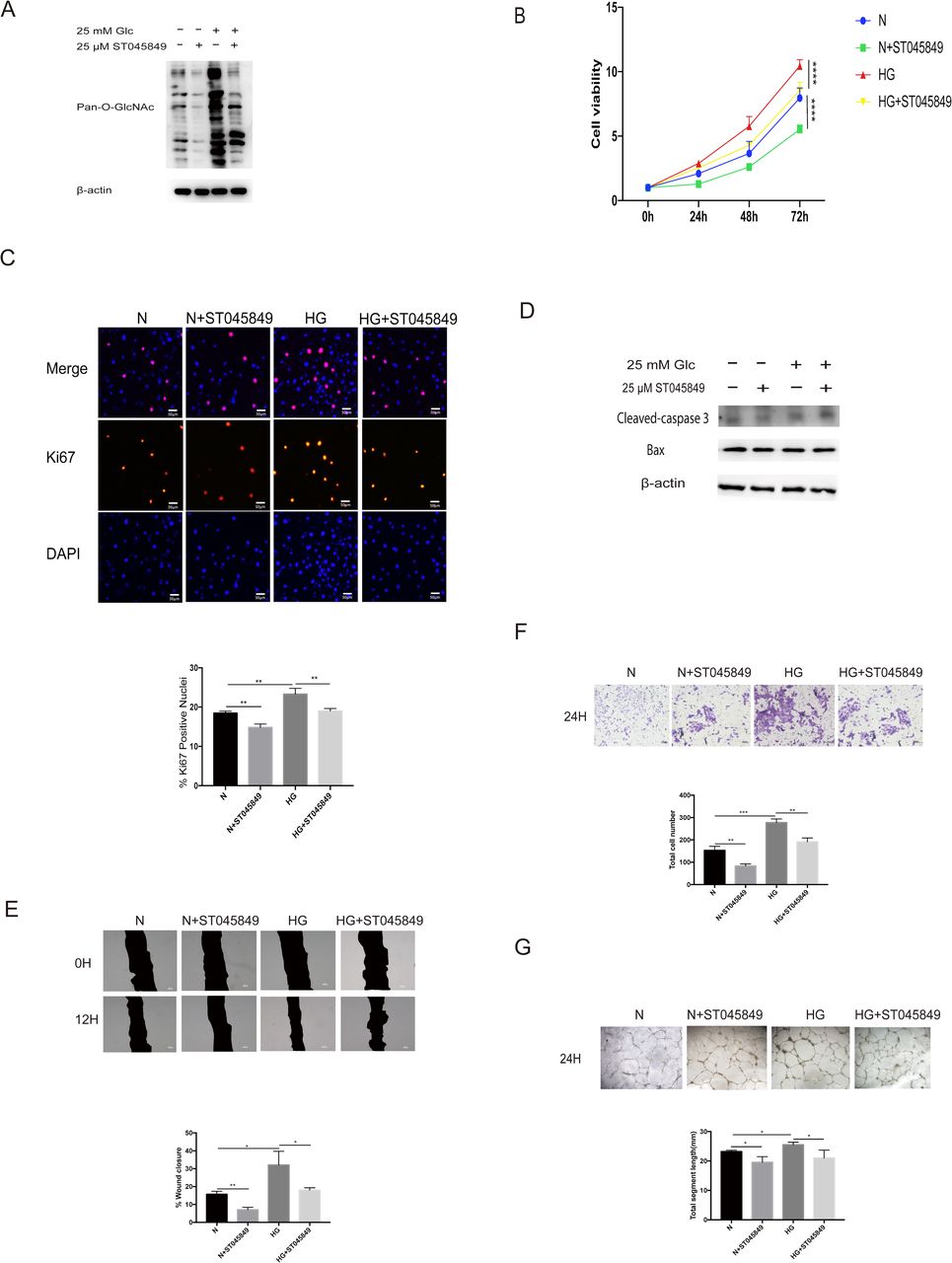
In order to assess the importance of O-GlcNAc modification on cell proliferation and migration, ST045849 was used to inhibit the OGT enzyme, that is, to inhibit the O-GlcNAc modification of the cells. First, we tested the inhibitory effect of ST045849. At the same glucose concentration, ST045849 could significantly inhibit the level of pan-cellular O-GlcNAc modification (figure 2A). No matter in normal glucose concentration or HG concentration, inhibition of O-GlcNAc modification can inhibit cell growth (figure 2B), and the decrease of Ki67-positive nuclei also indicates that inhibiting O-GlcNAc modification can inhibit the proliferation of cells (figure 2C). In this part of the experiment, we still choose scratch-wound migration assay and Transwell migration assay test to detect cell migration. Before that, we also excluded the effect of apoptosis on migration detection using western blot analysis, there was no change in apoptosis among the groups (figure 2D). In HRMECs, 25 µM of ST045849 could effectively reduce wound healing area and total number of transmembrane cells (figure 2E,F), which means that the level of O-GlcNAc modification is very important for cell migration. Similarly, we examined the effect of ST045849 on tube formation. We found that ST045849 could significantly inhibit the formation of tube at the same glucose concentration (figure 2G).
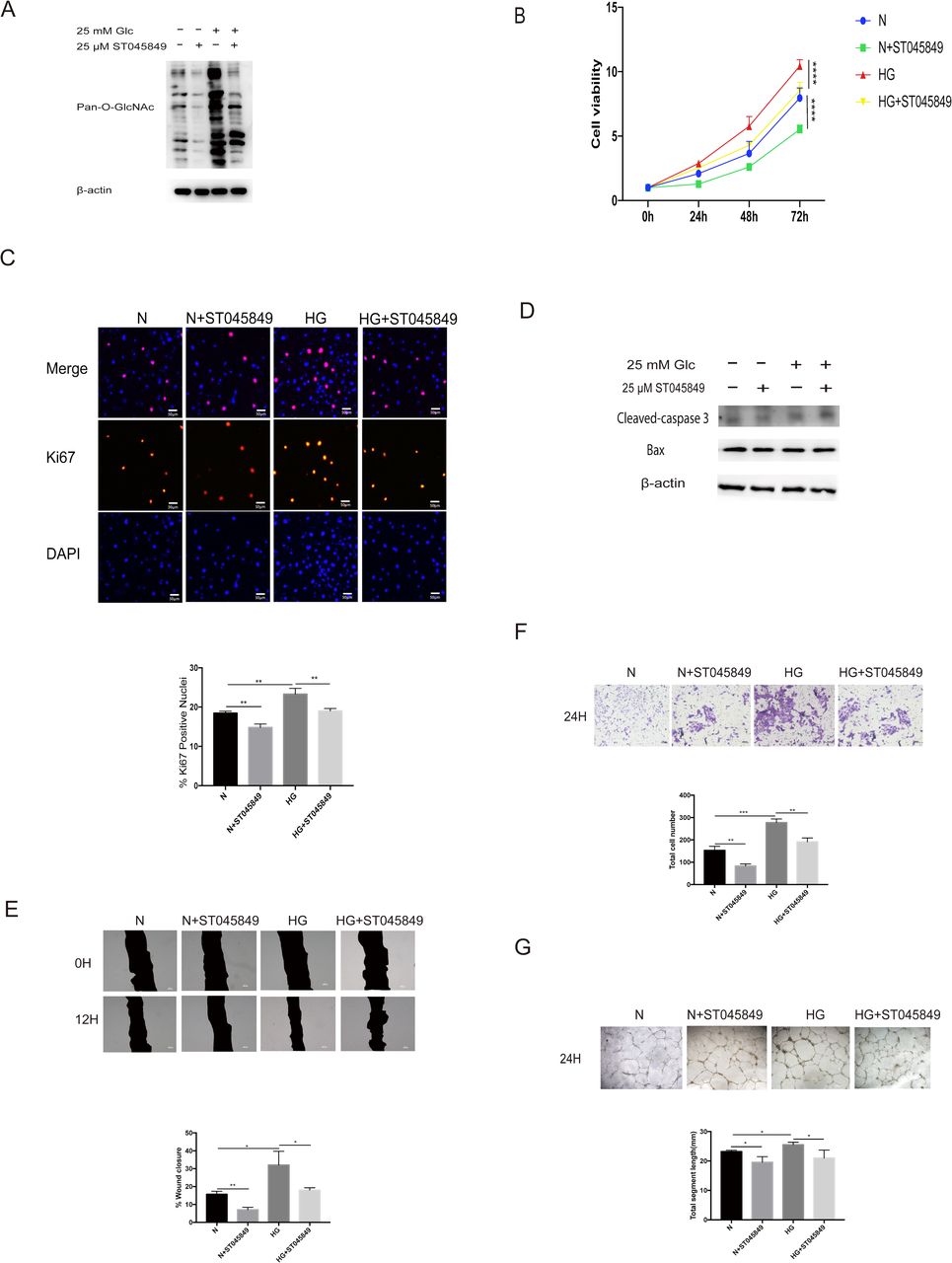
O-linked N-acetylglucosamine (O-GlcNAc) modification is critical for the proliferation and migration of human retinal microvascular endothelial cells (HRMECs). Cells were treated with 5.5 mM of glucose (normal (N)) or 5.5 mM+25 µM of ST045849 (N+ST045849) or 25 mM of glucose (high glucose (HG)) or 25 mM of glucose+25 µM of ST045849 (HG+ST045849). (A) The protein lysates were collected and processed by western blot analysis. Western blot analysis was performed using antibodies specific for O-GlcNAc modification and β-actin. (B) Cell growth was determined by cell counting kit-8 (CCK-8), the graphs present the analysis of cell viability. (C) Cell proliferation was determined by Ki67 staining, the graphs present the analysis of ki67-positive rate. Scale bar=50 µm. (D) The apoptosis was detected by western blot analysis. Western blot analysis was performed using antibodies specific for cleaved-caspase 3, Bax, and β-actin. (E) Scratch-wound assay using HRMECs at 0 (top) and 12 hours (bottom). The black area represents the scratch area. The graphs represent the scratch healing rate. Scale bar=100 µm. (F) The migratory ability in HRMECs were detected by Transwell assay. The purple dots represent the cells that completed the vertical migration (passed through the membrane). The graphs represent the numbers of cells that passed through the membrane. Scale bar=100 µm. (G) Tube formation ability was detected by tube-forming assay. Cells were re-incubated on Matrigel. The graphs represent total segment length; n=3, independent experiments. Scale bar=100 µm. *P<0.05, **p<0.01, ***p<0.001, ****p<0.0001.
Result 3: RUNX1 mediates the glucose and O-GlcNAc-driven proliferation and migration of HRMECs
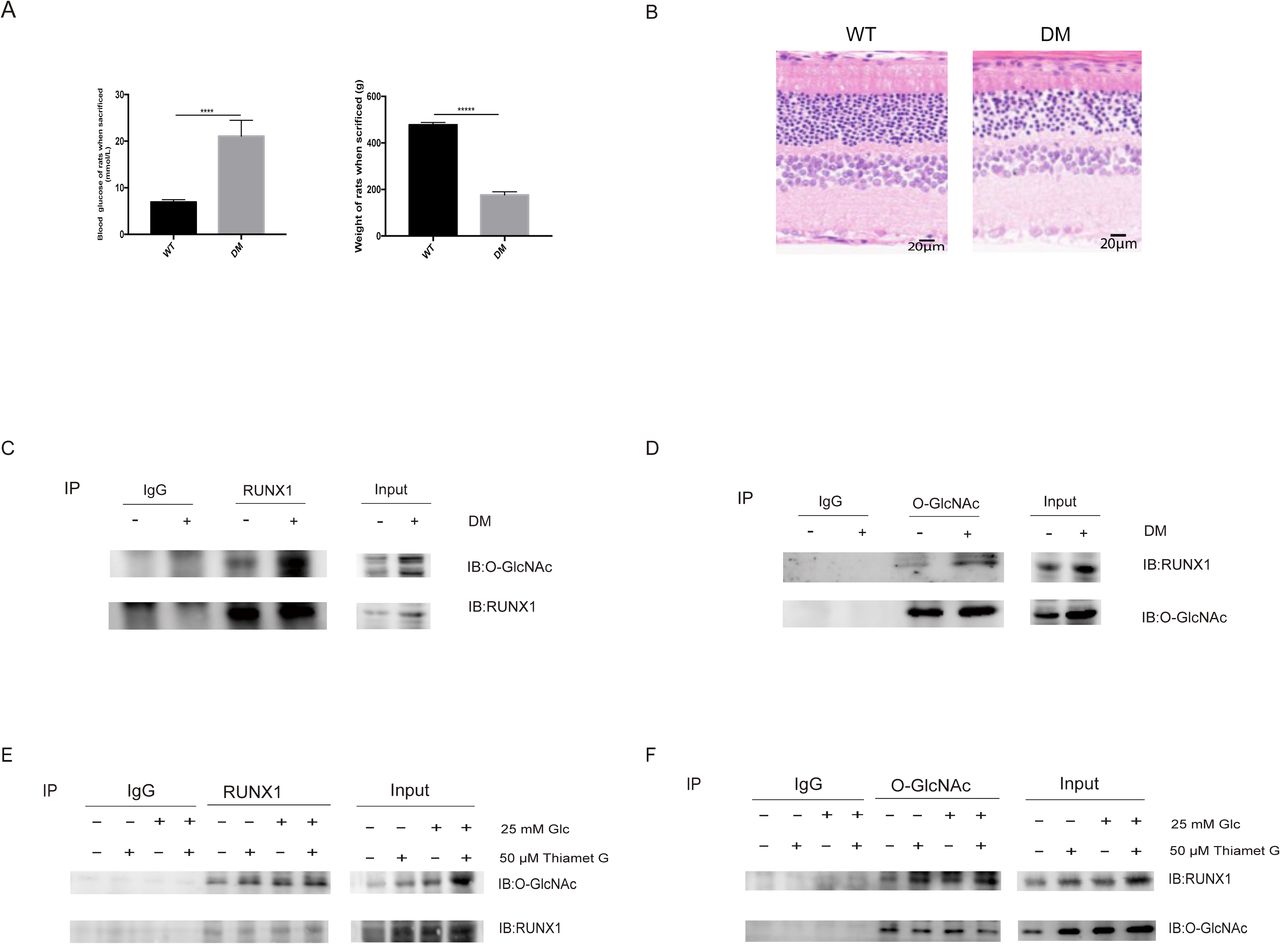
Previous studies have revealed that the inhibition of RUNX1 can significantly inhibit the formation of retinal neovascularization in hypoxia-induced retinal neovascularization mouse model. The knockdown of RUNX1 in HRMECs can reduce the ability of cells to form tubes,23 indicating that RUNX1 has a clear role in promoting angiogenesis, but its mechanism remains unknown. In order to evaluate the role of RUNX1 in the HG and O-GlcNAc-mediated induction of cell proliferation and migration, HRMECs depleted of RUNX1 by siRNA were used. The control cells expressed a scramble siRNA sequence. The RUNX1 knockdown was verified by western blot analysis prior to the experiments. siRUNX1 can effectively knockdown RUNX1 protein in cells at normal glucose concentration or HG concentration, whether using Thiamet G or not, and siRUNX1 did not significantly interfere with the pan-cellular O-GlcNAc modification, compared with cells expressed a scramble siRNA sequence (figure 3A). siRUNX1 significantly slowed down cell growth, whether caused by HG or high O-GlcNAc modification level (figure 3B). HRMECs transfected with RUNX1 siRNA had fewer Ki67-positive cells compared with cells transfected with a scramble siRNA sequence 24 hours after transfection, indicating RUNX1 also contributes to HRMECs proliferation, whether it is caused by HG or high level of O-GlcNAc modification (figure 3C). To investigate the potential pro-angiogenic effect of RUNX1, scratch healing ability, penetration ability, and tube-forming ability were tested. Of course, before that, we also detected the apoptosis that may affect the migration and tube-formation abilities. The results showed that there was no significant difference in apoptosis among the groups (figure 3D). As shown in figure 3E,F, siRUNX1 can significantly slow down wound healing and reduce the total number of cells passing through the membrane caused by HG or high level of O-GlcNAc modification, indicating that the presence of RUNX1 is very important for cell migration. Figure 3G shows that knockdown of RUNX1 can significantly reduce the tube formation. Next, we explored the possible mechanism of RUNX1. STZ is a pancreatic islet β-cell-cytotoxic antibiotic that can highly and selectively destroy pancreatic islet β cells, and induce hypoinsulinemia and hyperglycemia.24 STZ-induced diabetes in rats is the most commonly used experimental model for investigating DR.25 We divided 40 rats into two groups, WT group and STZ-induced DM group, 20 rats in each group. Figure 4A shows the differences of blood glucose and body weight between the two groups. The blood glucose of DM rats is significantly higher than that of WT rats, and the body weight of DM rats is significantly lower than that of WT rats. Before the sacrifice, the retina structure of rats was obviously disordered, which indicated that they had suffered from DR (figure 4B). In order to minimize the effect of individual difference, we mixed the retinal tissues of five rats as a sample, three samples in each group. RUNX1 was immunoprecipitated from samples. The western blot analysis revealed that the RUNX1 protein can be modified by O-GlcNAc, and that a hyperglycemic environment can promote its O-GlcNAc modification (figure 4C,D). Then we tested the O-GlcNAc modification status of RUNX1 under 5.5 mM and 25 mM glucose concentrations, with and without Thiamet G, as shown in figure 4E,F, both HG condition and treatment of cells with Thiamet G could increase the O-GlcNAc modification of RUNX1.
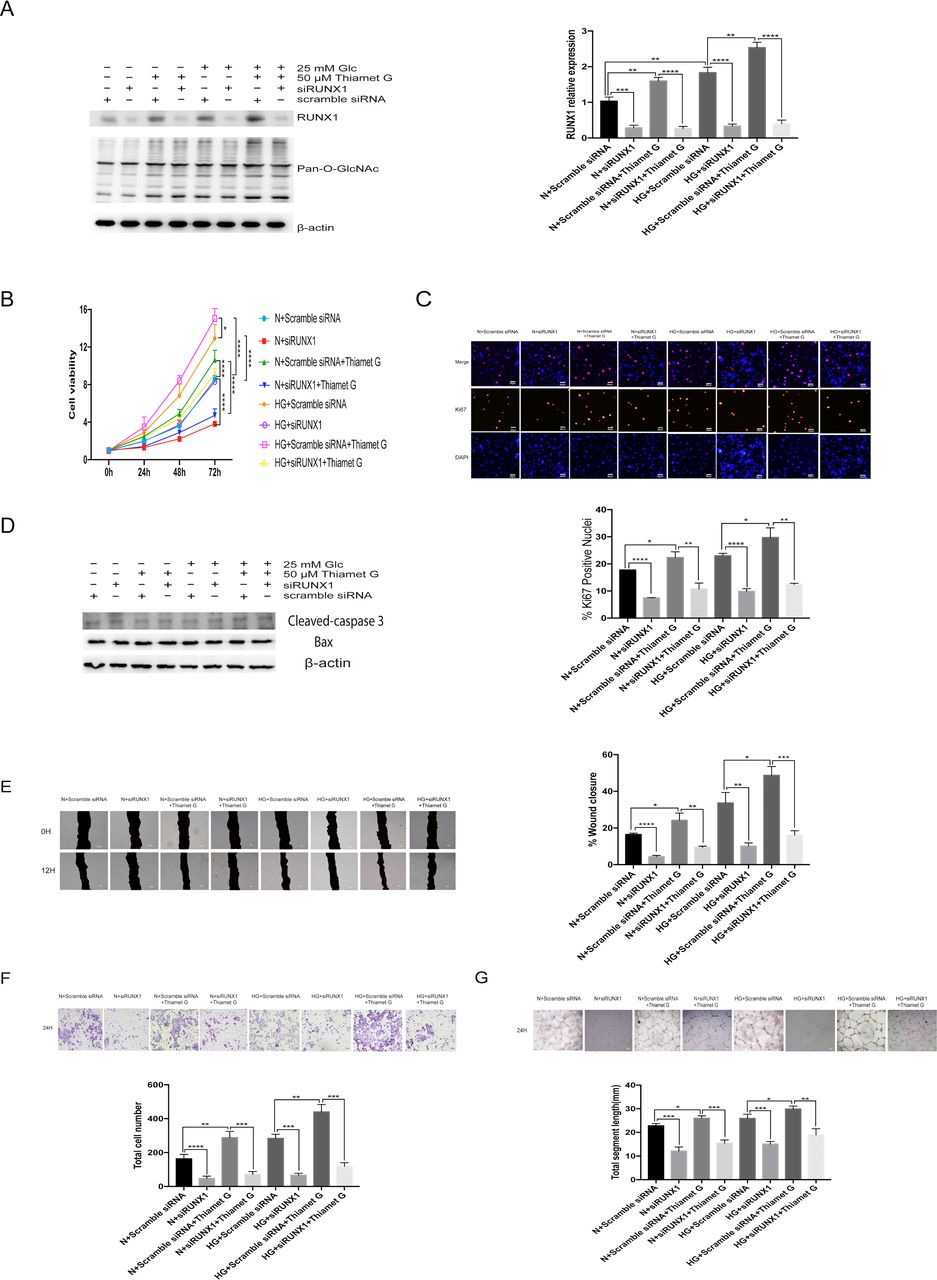
Runt-related transcription factor 1 (RUNX1) mediates the glucose and O-linked N-acetylglucosamine (O-GlcNAc)-driven proliferation and migration of human retinal microvascular endothelial cells (HRMECs). Cells were treated with 5.5 mM of glucose+scramble small interfering RNA (siRNA) (normal (N)+scramble siRNA) or 5.5 mM of glucose+siRUNX1 (N+siRUNX1) or 5.5 mM of glucose+scramble siRNA+50 µM of Thiamet G (N+scramble siRNA+Thiamet G) or 5.5 mM of glucose+siRUNX1+50 µM of Thiamet G (N+siRUNX1+Thiamet G) or 25 mM of glucose+scramble siRNA (high glucose (HG)+scramble siRNA) or 25 mM of glucose+siRUNX1 (HG+siRUNX1) or 25 mM of glucose+scramble siRNA+50 µM of Thiamet G (HG+scramble siRNA+Thiamet G) or 25 mM of glucose+siRUNX1+50 µM of Thiamet G (HG+siRUNX1+Thiamet G). (A) The protein lysates were collected and processed by western blot analysis. Western blot analysis was performed using antibodies specific for O-GlcNAc modification and β-actin. The graphs present the gray value analysis of RUNX1 band. (B) Cell growth was determined by cell counting kit-8 (CCK-8), the graphs present the analysis of cell viability. (C) Cell proliferation was determined by Ki67 staining, the graphs present the analysis of Ki67-positive rate. Scale bar=50 µm. (D) The apoptosis was detected by western blot analysis. Western blot analysis was performed using antibodies specific for cleaved-caspase 3, Bax, and β-actin. (E) Scratch-wound assay using HRMECs at 0 (top) and 12 hours (bottom). The black area represents the scratch area. The graphs represent the scratch healing rate. Scale bar=100 µm. (F) The migratory ability in HRMECs were detected by Transwell assay. The purple dots represent the cells that completed the vertical migration (passed through the membrane). The graphs represent the numbers of cells that passed through the membrane. Scale bar=100 µm. (G) Tube formation ability was detected by tube-forming assay. Cells were re-incubated on Matrigel. The graphs represent total segment length; n=3, independent experiments. Scale bar=100 µm. *P<0.05, **p<0.01, ***p<0.001, ****p<0.0001.
{kind=link}
{kind=link}
{kind=link}
{kind=link}
High glucose (HG) environment and treatment of cells with Thiamet G can promote the O-linked N-acetylglucosamine (O-GlcNAc) modification of Runt-related transcription factor 1 (RUNX1). Rats with diabetes mellitus (DM) were induced by intraperitoneal injection of streptozotocin (STZ). Cells were treated with 5.5 mM of glucose (normal (N)) or 5.5 mM+50 µM of Thiamet G (N+Thiamet G) or 25 mM of glucose (HG) or 25 mM of glucose+50 µM of Thiamet G (HG+Thiamet G) for 48 hours. (A) Blood glucose and body weight of wild-type (WT) and DM rats. (B) H&E staning of WT and DM rat retina tissues. Scale bar=20 µm. (C) O-GlcNAc modification of RUNX1 in retinas was detected by immunoprecipitation (IP), RUNX1, and IgG was immunoprecipitated from the retinas of WT and DM rats (IP), western blot analysis was performed using antibodies specific for O-GlcNAc modification and RUNX1 (IB), IgG, and Input were control groups. (D) O-GlcNAc modification of RUNX1 in retinas was detected by IP, O-GlcNAc, and IgG was immunoprecipitated from the retinas of WT and WT rats (IP), western blot analysis was performed using antibodies specific for O-GlcNAc modification and RUNX1 (IB), IgG, and Input were control groups. (E) O-GlcNAc modification of RUNX1 in HRMECs was detected by IP, RUNX1, and IgG was immunoprecipitated from HRMECs (IP), western blot analysis was performed using antibodies specific for O-GlcNAc modification and RUNX1 (IB), IgG, and Input were control groups (IB), IgG, and Input were control groups. (F) O-GlcNAc modification of RUNX1 in HRMECs was detected by IP, O-GlcNAc, and IgG was immunoprecipitated from HRMECs, western blot analysis was performed using antibodies specific for O-GlcNAc modification and RUNX1 (IB), IgG, and Input were control groups; n=3, independent experiments.
Discussion
The findings of this study reveal that hyperglycemia directly affects the function of HRMECs. Furthermore, O-GlcNAcylation is critical for the proliferation and migration of HRMECs, and RUNX1 may take part in this mechanism.
Previous studies have shown that O-GlcNAcylation is involved in the pathological changes of many diseases, such as diabetes, Alzheimer’s disease, various cancers, and AIDS.26–28 O-GlcNAc modification affects the stability and activity of many transcription factors, such as p53, STAT5, PDX-1, and FOXO129–32 by modifying these transcription factors and exhibits various effects on diseases. O-GlcNAcylation can reduce the apoptosis of coronary endothelial cells and improve the performance of diabetic heart disease through p53 and histone deacetylase 4.33 34 O-GlcNAc modification can also regulate tumor growth and cancer cell proliferation and migration through AMPK, pyruvate kinase M2, and β-catenin.35–37 In DR, the protective effect of increased O-GlcNAc modification against reactive oxygen species has been reported in human retinal endothelial cells.38 Overexpression of hyperglycemia-induced intracellular adhesion molecule 1 is linked to the O-GlcNAc modification of transcription factor Sp1 in human umbilical vein endothelial cells and rat retinal capillary endothelial cells.39 The increase in O-GlcNAcylation of Sp1 is associated with the upregulated VEGF-A production in the retinal endothelium and pigment epithelium.19 These results suggest that O-GlcNAcylation may affect the function of endothelial cells and diabetic neovascularization. The present study provides proof and supplements this conjecture.
Previous studies have shown that RUNX1 targets many signaling pathways, such as transforming growth factor (TGF)-β signaling pathway and Wnt signaling pathway.40 41 TGF-β can regulate the differentiation, proliferation, apoptosis, and migration of many kinds of cells, and also influence the formation of blood vessels.40 The Wnt signaling pathway takes part in cell proliferation, migration, and apoptosis.41 Cell proliferation and migration are the processes of angiogenesis. Recent studies have shown that RUNX1 can affect the proliferation and migration of retinal endothelial cells and block endothelial cell tube formation and aberrant angiogenesis in an oxygen-induced retinopathy model.23 However, the mechanism has not been demonstrated. In this study, we found that HG could increase RUNX1 expression, this increase was achieved at the transcriptional level because it could not be maintained after siRUNX1 transfection.
Increasing the level of O-GlcNAc modification by treating cells with Thiamet G can also increase the RUNX1 expression, this increase is ultimately achieved by affecting the transcription level because it could not be maintained after siRUNX1 transfection. Possibly, O-GlcNAc modification can affect other related proteins and thus affect the expression of RUNX1. Whether HG can increase the expression of RUNX1 independently from O-GlcNAcylation is still not clear because HG concentration can increase O-GlcNAc modification by itself, and also can cause many other changes, and the relationship between them is very complex. Recently, it was found that O-GlcNAc can modify RUNX1, and the modification is enhanced in a HG environment. Treatment of cells with Thiamet G also could increase the O-GlcNAcylation. This is consistent with the function of RUNX1 under HG and high O-GlcNAcylation conditions. Therefore, we may suggest that the function of RUNX1 is related to the level of its O-GlcNAc modification.
There are some limitations to the present study. The O-GlcNAc site of RUNX1 was not obtained. Through the specific blocking of the O-GlcNAcylation of RUNX1, its role can be more precisely determined. Furthermore, the impairment of the surrounding cells and microenvironment may have adverse effects on retinal EC as the DR progresses, and this is coupled with both hyperglycemia and increased O-GlcNAcylation. However, further studies on retinal cells and in animal tissues are needed.
In conclusion, the present study reveals that HG condition directly affects retinal EC function, and O-GlcNAcylation is critical for the proliferation and migration of HRMECs. Moreover, RUNX1 may take part in this mechanism, and the function of RUNX1 is related to its O-GlcNAc modification level.
Data availability statement
All data relevant to the study are included in the article.
Ethics statements
Ethics approval
The experimental protocols used in the present study followed the guidelines established by the ARVO Statement for the Use of Animals in Ophthalmic and Vision Research, and were approved by the Ethics Committee of Shanghai General Hospital, Shanghai Jiaotong University, Shanghai, China.
References
Footnotes
Contributors Conception and design: XX and KL. Data acquisition and analysis: XX and KL. Data interpretation, drafting of the manuscript and approval of the final version: all authors.
Funding This study was supported by National Key R&D Program of China (grant no: 2019YFC0840607), the National Natural Science Foundation (grant no: 81870667).
Competing interests None declared.
Provenance and peer review Not commissioned; externally peer reviewed.